
Investigating changes in the post-transcriptional pattern of VEGF and CD146 genes carrying miR-573 in breast cancer cells treated with tamoxifen
Zkoumání změn posttranskripčního úprav genů VEGF a CD146 nesoucích miR-573 v buňkách karcinomu prsu ošetřených tamoxifenem
Východiska: Nejnovější výzkumy týkající se podílu microRNA (miRNA) na nádorové angiogenezi a onkogenních účinků miRNA ukazují na jejich potenciální roli při angiogenezi karcinomu prsu. Exozomy získané z nádoru jsou považovány za bohatý zdroj miRNA, které regulují funkce ostatních buněk v nádorovém mikroprostředí, vč. endoteliálních buněk cév. Tato studie analyzuje účinek chemoterapie tamoxifenem na expresi klíčové miRNA, miR-573, která se účastní angiogeneze v exozomech nádoru a působí jako regulační spojka mezi miRNA a genem CD146, který je součástí dráhy vaskulárního endoteliálního růstového faktoru (vascular endothelial growth factor – VEGF).
Materiál a metody: Byly zakoupeny buňky karcinomu prsu MCF-7 a kultivovány v kompletním médiu. Tyto buňky byly ošetřeny tamoxifenem a z média byly následně exrahovány jejich exozomy. Byla izolována RNA exozomů a pomocí metody polymerázové řetězové reakce (PCR) v reálném čase byla vyhodnocena exprese genů miR-573, VEGF a CD146 v exozomech.
Výsledky: Výsledky této studie ukázaly, že ošeření buněk tamoxifenem zvýšilo expresi miR-573 v exozomech získaných z nádorových buněk MCF-7. Exprese genů CD146 a VEGF v exozomech buněk ošetřených léčivem měla klesající tendenci.
Závěr: Výsledky tohoto pokusu prokázaly, že ošetření nádorových buněk tamoxifenem snižuje expresi genů VEGF and CD146 zvýšením miR-573. Dochází tedy k omezení angiogeneze a tím k protinádorovému působení léčiva.
Klíčová slova:
karcinom prsu – nádorová angiogeneze – exozom – tamoxifen – miR-573
Authors:
M. Abyar; M. Kavousi; F. Jamshidian
Authors place of work:
Department of Biology, East Tehran Branch, Islamic Azad University, Tehran, Iran
Published in the journal:
Klin Onkol 2023; 36(6): 447-452
Category:
Původní práce
doi:
https://doi.org/10.48095/ccko2023447
Summary
Background: Recent developments regarding the contribution of microRNAs (miRNAs) to tumor angiogenesis and the oncogenic effects of miRNAs point to their potential role in breast cancer angiogenesis. Tumor-derived exosomes are considered a rich source of miRNAs that can regulate the function of other cells in the tumor microenvironment, including vascular endothelial cells. This study analyzes the effect of tamoxifen chemotherapy on the expression of a key miRNA, miR-573, involved in the angiogenesis of the tumor exosomes and introduces a regulatory link between this miRNA and the CD146 gene associated with the vascular endothelial growth factor (VEGF) messaging pathway.
Materials and methods: MCF-7 breast cancer cells were purchased and cultured in a complete culture medium. These cells were treated with tamoxifen and then their exosomes were extracted from the culture medium. The RNAs of the exosomes were isolated and the expression of miR-573, VEGF, and CD146 genes in the exosomes was investigated using the real-time polymerase chain reaction (PCR) method.
Results: The results of this study showed that tamoxifen treatment increased the expression of miR-573 in exosomes derived from MCF-7 cancer cells. The expression of CD146 and VEGF genes in drug-treated cell exosomes had a downward pattern.
Conclusion: The results of this experiment demonstrated that the treatment of breast cancer cells with tamoxifen reduces the expression of VEGF and CD146 by increasing miR-573. Thus, angiogenesis is reduced and, therefore, its anti-tumor effects are applied.
Keywords:
breast cancer – tamoxifen – tumor angiogenesis – exosome – miR-573
Introduction
Breast cancer occurs as a result of the abnormal growth of cells in the breast. That growth pattern stops at a certain stage in benign tumors, whereas the malignancy development continues uncontrollably in malignant tumors that if left untreated, affect and disable all parts of the body [1]. Tamoxifen is a drug used to reduce or stop the effects of the female hormone, estrogen, in the body and is one of the drugs used in breast cancer treatment [2]. MicroRNAs (miRNAs) are a new class of endogenous non-coding RNA molecules that regulate gene expression at the post-transcriptional level by mRNA degradation and/or translation inhibition. There exist more than 2,000 miRNA-encoding genes in the human genome, which make up about one percent of the whole genome [3]. The essential role of miRNA in the processes of carcinogenesis and oncogenesis is obvious. Available evidence suggests that abnormal levels of expression in some specific miRNAs can cause misalignment of cancer cells. MiRNA molecules can play an important role in disease control at the biological stages. These miRNAs can act as oncogenes or tumor suppressor genes. The study of the expression of total genome miRNAs in recent years has shown more detailed information about the intervening role of these small regulatory molecules in human cancers [4].
Exosomes are small vesicles (50–100 nm) that have recently been known as intercellular communication linkers. Exosomes can transfer lipids, proteins, mRNAs, and miRNAs to the receptor cell when moving to other cells. Cancer cells have been shown to secrete far more exosomes than normal cells, thus transmitting tumor-dependent signaling molecules to surrounding cells [5].
MiRNAs are among the molecules being transferred by exosomes and regulate most of the cell‘s biological functions. They regulate translation and transcription programs and coordinate pathological as well as physiological processes such as development, cell differentiation, proliferation, apoptosis, and tumor growth [6]. MiRNAs act as small guide molecules in RNA silencing and negatively regulate the expression of various genes, both at the miRNA level and the protein level, by degrading the target miRNA and silencing the translation. One of the recent developments in this field of study is the understanding of their role in oncogenesis. The number of miRNA genes is increasing, and changes in the level of miRNAs play a role in the initiation, progression, and metastasis of various tumors. Biochemical and genetic evidence confirms the role of miRNAs in tumor development. The abnormal expression of miRNAs in cancer cells is a well-known phenomenon, the cause of which is yet unidentified [7]. The exact mechanism by which tumor cell-derived exosomes affect their constituent microenvironmental cells, including endothelial cells, has not yet been determined, and the biological function of exosomal miRNAs in receptor cells is not well understood. Therefore, this study investigates the effect of tamoxifen chemotherapy drug on the expression of a key miRNA involved in angiogenesis, miR-573, in tumor exosomes and introduces a regulatory relationship between this miRNA and CD146 gene, associated with vascular endothelial growth factor (VEGF) messaging pathway.

Materials and methods
Cell preparation and culture
The MCF-7 breast tumor cell line with NCBI CODE 135 and HUVEC cell line were both purchased from the Iranian Genetic Resources Center. Subsequently, the cells were cultured in a full culture medium containing FBS and trypsin (GibCo). The flasks were kept inside an incubator with CO2 saturation rate 5% and a temperature of 37 °C.
Tamoxifen treatment
Based on the results of various experiments and resource reviews [8], tamoxifen IC50 was a concentration between 54 μM (equivalent to 20 μg/mL) and 68 μM (equivalent to 25 μg/mL). Hence, the concentration of 60 μM (as a concentration close to IC50) was considered for the treatment in this study.
Exosome isolation
Exosomes were isolated from the culture medium of MCF-7 cells. For this purpose, the culture medium was first centrifuged at 3,000 g for 10 minutes to separate the dead cells from the culture medium. Exoquick solution (System Biosciences) was then added to the supernatant. Vortex binds the exoquick solution to exosomes. Then it was incubated for 20 minutes at 4 °C. The centrifugation was afterwards performed at a speed of 1,500 g for 15 minutes. 50μL PBS was then added to the solution and pipetting was performed. The content was transferred to a microtube and placed on a heater for 5 minutes at 70°C and then at −20 °C. Exosome isolation was performed from the supernatants of both treated and control cell groups, confirmation of which was done by microscopic examination, digital electron microscope (Digital FESEM, KYKY-EM3200, China), evaluating the size and morphology of the exosomes.
Exosome labeling
PKH26 (PKH26 Red fluorescent – Mini26-1, Sigma) was used to label the exosomes. One mL of the exosome was transferred to a microtube, and 1 mL of diluent C was added to it and gently pipetted. From the diluted paint PKH26, 1 mL was added into the microtube which contained the exosome, and then incubated at room temperature for 5 min. To stop the labeling, 2 mL of diluent C was again added into the microtube and placed at room temperature for 5 more minutes. Then the exoquick solution was added with a ratio of 3 : 1 and kept at 4 °C for 20 minutes. Later, the solution was centrifuged for 20 minutes at 15,000 g. After centrifugation, the top solution was removed and the exosome-containing precipitate was dissolved in 30 μB PBS. The endothelial cell was then treated with stained exosomes, followed by incubation at room temperature for 12 hours. Due to the presence of miRNAs in exosomes, the exosomal transmission of miR-206 to fibroblast cells was investigated. In this regard, fibroblast cells were treated by 100 µg/mL tumor exosomes and 50 µg/mL alpha-amanitine. The rate of miR-206 transferring to fibroblast cells was evaluated at 0, 24, and 48 hours post-treatment. The reason for using alpha-amanitine is to inhibit any transcription activity that can be induced by exosomal treatment in cells.
RNA isolation
RNA isolation from cells and exosomes was performed using the Trizol (Invitrogen) agent. After centrifuging the cells and adding 50 μL of Trizol solution, 15 μL of chloroform was added to it and incubated at room temperature for 2–3 minutes. After centrifugation at 12,000 g, the upper phase was removed and isopropanol was added and then the whole solution was incubated on ice for 10 minutes. The centrifugation was performed another time at 12,000 g for 10 minutes. Subsequently, 1 mL of ethanol 75% was added to the residual precipitate, and centrifugation was performed again at 7,500 g at 4 °C for 5 min. The RNA sample was stored in a freezer at −80 °C. The quantitative analysis of the isolated RNA was done using spectrophotometry (NanoDrop Technologies, Wilmington, DE, USA).
Investigating the expression level of CD146, VEGF, and miR-573
cDNA was generated using the 1st strand cDNA Synthesis kit (Thermo Fisher Scientific), Oligo dT, and random hexamer primers. In order to perform the real-time PCR reaction, the forward and reverse primers (Tab. 1), cDNA, DEPC water, Mastermix, and CybrGreen dye were all added and the solution was placed in a thermocycler. Data analysis was performed using the threshold cycle comparison method according to the table below. Normalization of changes in miRNA expression level was done in comparison with the endogenous U6 snRNA expression level, while the expression level of other genes was compared with GAPDH. Data analysis was performed using ABI step one Real-Time PCR Software v2.0.2 (Applied Biosystems, UK), Excel software (Microsoft, 2010), and GraphPad software.
Statistical analysis
Statistical analysis of the obtained data was performed using the SPSS v.20 software, Anova test and Tukey follow-up test. P < 0.05 was considered significantly meaningful in this study.
Results
Exosome isolation confirmation
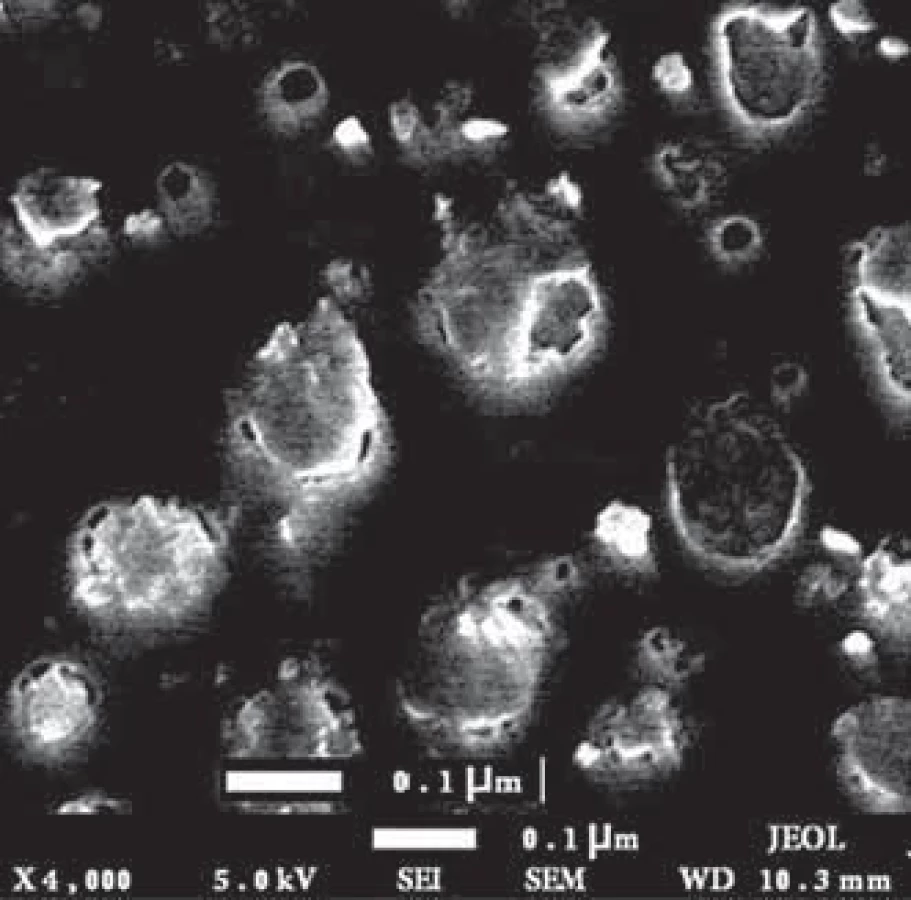
The size and morphology of the exo - somes were evaluated by electron microscopy. The results showed that the isolated exosomes had a spherical appearance with a size range of 30–150 nm (Fig. 1).
Measurement of endothelial cells‘ ability to absorb PKH26 fluorescent-labeled tumor exosomes
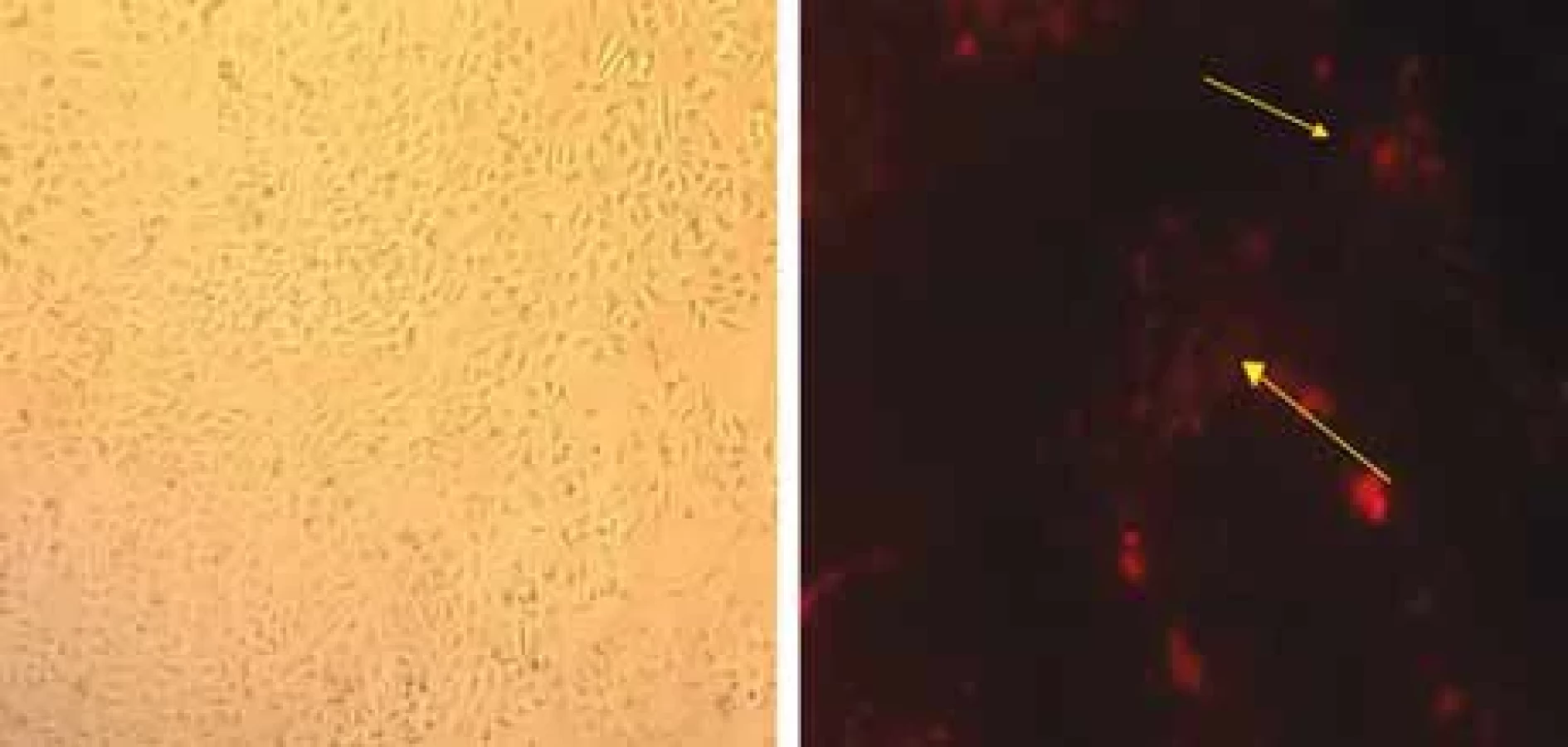
In order to determine the ability of endothelial cells to absorb tumor exosomes, HUVEC endothelial cells were treated by PKH26-labeled fluorescent colored exosomes. They were then incubated for 12 hours. Fluorescent microscopic analysis showed that PKH26 fluorescent-labeled exosomes are located in the cytoplasm of endothelial cells (Fig. 2). This indicates the ability of HUVECs to absorb endosomes derived from breast tumor cells.
Examination of miR-573 expression level in exosomes isolated from MCF-7 breast cancer cells
According to Graph 1, the miR-573 expression level in tumor cell-derived exosomes treated with tamoxifen has a higher expression than exosomes derived from the control cell group. This result suggests that tamoxifen treatment may alter the expression profile of miRNAs which have tumor suppressor activities such as miR-573 in tumor cell-derived exosomes. The expression level of miR-573 being treated by tamoxifen has up-regulation.
Transferring miR-573 from MCF-7 breast tumor cell-line to HUVEC endothelial cells
Initially, the exosomal transfer of miR-573 to endothelial cells was investigated due to the presence of miRNAs in the exosomes. For this reason, HUVEC endothelial cells were first treated by 100 µg/mL alpha-amanitine, and then the rate of miR-573 transfer to endothelial cells was assessed at 0, 24, and 48 hours post-treatment. The reason lying behind using alpha-amanitine is to inhibit any transcription activity that can be induced by exogenous exosomes in the endothelial cells. As it is seen in Graph 2, the level of miR-573 transcripts in endothelial cells treated by exosomes and alpha-amanitine increased significantly over time.
Effects of breast tumor cells-derived exosomes on the expression level of CD146 and VEGF genes: tumor-derived exosomes that have been left untreated induced the expression level of CD146 and VEGF genes time-dependently. Much to our surprise, in contrast, tamoxifen-treated tumor exosomes reduced CD146 and VEGF expression levels over time. Therefore, given the greater presence of miR-573 in tamoxifen-treated tumor exosomes, it is comprehended that the miR-573 exosomal transfer reduces the expression level of CD146 and VEGF in endothelial cells after 24 and 48 hours significantly and time-dependently. Therefore, it is assumed that the exosomal transfer of miR-573, which is a tumor suppressor, from the mammary tumor cells being treated by tamoxifen may have an inhibitory effect on angiogenesis through decreasing the activity and expression level of CD146 and VEGF pro-angiogenic genes (Graphs 3,4).




Discussion
Cancer cells seek to activate regulatory pathways for survival and development, as well as to obtain the metabolites and growth factors they need [9]. Since angiogenesis is developed by microenvironmental changes in cancer cells through tumor exosomes, it appears that inhibiting angiogenesis is mediated by the anti-angiogenic chemotherapy drug, tamoxifen, through modulating related miRNAs‘ expression. Therefore, it is assumed that tumor cell exosomes should be targeted to inhibit angiogenesis [10]. Tumor-derived exosomes are considered as a rich source of miRNAs that could regulate the function of other cells in the tumor microenvironment, including vascular endothelial cells. However, the exact mechanisms by which tumor cell-derived exosomes affect their microenvironmental cells, as well as the biological function of exosomal miRNAs in cells, have not been well established [11]. Therefore, this study evaluated the effect of one of the chemotherapy drugs, tamoxifen, on the expression level of a key miRNA involved in angiogenesis, miR-573, and tried to introduce a regulatory relationship between this miRNA and CD146 gene associated with the VEGF messaging pathway. The results showed that the expression of miR-537 in the exosomes derived cells treated by tamoxifen increased significantly and time-dependently. On the other hand, the expression of CD146 and VEGF genes in drug-treated cell exosomes had a downward pattern.
In 2015, Tahakashi et al. investigated the breast tumor-derived miRNAs and introduced these miRNAs as a diagnostic biomarker in the treatment of breast cancer [12]. The result of the presented study, in line with the research of Tahakashi et al., confirmed the role of miR-573 as a diagnostic biomarker in breast cancer. In 2015, Hanafen et al. concluded that the use of the anti-cancer drug docosahexaenoic acid (DHA) changes the exosomal secretions of breast cancer cells. They isolated MCF7 and MDA-MB-231 breast cancer cell exosomes after treatment with DHA and observed an increase in the secretion of exosomes and the content of miRNAs of DHA-treated exosomes. The expression level of miRNA-83 in MCF7-treated exosomes had changed. The group concluded that DHA alters the exosomal secretions of breast cancer cells and their miRNA content, thereby inhibiting angiogenesis [13]. Although the chemotherapy drug used in the present study, tamoxifen, is different from that of Hanafen et al., the results are in line with each other in terms of diminishing the expression level of genes targeting the angiogenesis and cancer development.
The CD146 molecule belongs to the family of immunoglobulins. Recent research suggests that CD146 is an endothelial biomarker that promotes angiogenesis. The antibody against CD146, called AA98, prevents the migration of endothelial cells and subsequently prohibit angiogenesis and the growth of human tumors, and plays an important role in cell migration, especially in invasive intermediate trophoblast cells. It has recently been discovered that the CD146 is a transferring inducer between the epithelium and the mesenchymal tissue, thus expanding the invasion of breast cancer cells. CD146 is known as a receptor on endothelial cells. Tumor secretions stimulate the dimerization of CD146 molecules, activation of p38 MAPKs / NF-kB, and activation of many gene-producing genes including VEGF, MMP-9, and IL-8. AA98, which is the anti-molecular antibody of CD146, interferes with this signaling. The association of CD146 with tyrosine kinase P59, phosphorylase P125, and paxillin promotes the migration of endothelial cells. CD146 promotes angiogenesis in both laboratory and somatic cell conditions [14].
Increasing the expression level of CD146 facilitates the endothelial response to the kinase SRC stimulated by VEGF or VEGF-induced SRC kinase family (SKF), p38 mitogen-activated protein kinase (MAPKs) and NF-kappaB activation, and subsequently increases the migration of endothelial cells and tube formation. CD146 is a co-receptor for VEGFR-2; therefore, suppression of CD146 by miR-329 impairs signaling and angiogenesis induced by VEGF [15].
The expression level of miR-573 decreases in various tumors and its re-expression in the cell prevents the growth and development of melanoma. Characteristics of rheumatoid arthritis, which is a chronic autoimmune disease, is angiogenesis and cartilage decay in which miR-573 has a role [16]. MiR-578 and miR-573 play a role in the process of angiogenesis in breast cancer associated with BRCA gene. Laboratory evaluations on HEK293, MCF-7, and SUM149PT cells to investigate the role of miR-578 and miR-573 have shown that these miRNAs have been suppressed in breast cancer associated with BRCA 1/2. MiR-578 and miR-573 are associated with the central connection, or focal adhesion, VEGF, and the signaling pathway of hypoxia inducible factor-1 (HIF-1). MiR-578 and miR-573 control angiogenesis in breast cancer associated with BRCA 1/2, and do so by regulating VEGF and inducing HIF-1 [17]. In line with this research, the present study showed that when the tamoxifen-treated exosomes were transferred to the endothelial cell line, the expression of VEGF gene had a decreasing pattern.
Conclusion
Based on the results of the present study on the MCF-7 breast cancer cell line, it was found that there was a significant relationship between 60 μM dose of tamoxifen and MCF-7 cell supernatant exosomes, in a way that the expression level of exosomal miR-573 showed a significant increase. In this study, the effect of tamoxifen-treated tumor-derived exosomes was evaluated on the HUVEC endothelial cell line to assess the process of angiogenesis and treatment of breast cancer in its early stages. Applying the experiment group to the endothelial cell line reduces the expression of CD146 gene in endothelial cells significantly over time. The second gene studied in this study was VEGF, the result of which was the same as CD146. Ultimately, this paper could conclude that the exosomal transfer of miR-573 to endothelial cells reduces the expression of genes responsible for the angiogenesis process, including VEGF and CD146, identification of which could mitigate the tumor process.
When breast cancer cells are treated with tamoxifen, the level of miR-573 which is a tumor suppressor miRNA in the exosomes of these cells, is much higher than in untreated cancer cells. In contrast, when tamoxifen-treated tumor-derived exosomes are transferred to endothelial cells, the expression level of CD146 and VEGF genes declines in a time-dependent manner. This achievement brings us to the idea that the therapeutic effect of tamoxifen on breast cancer lies behind its relationship with the expression level of exosomal miRNAs that regulate the cancer microenvironment.
Acknowledgments
This study was conducted at the cellular and molecular laboratory of Islamic Azad University, thus appreciating the spiritual support of these centers is admitted.
Availability of data and materials
The data used in this study are available from the corresponding author upon request.
Zdroje
1. Albeshan SM, Mackey MG, Hossain SZ et al. Breast cancer epidemiology in gulf cooperation council countries: a regional and international comparison. Clin Breast Cancer 2018; 18 (3): e381–e392. doi: 10.1016/ j.clbc.2017.07.006.
2. Rondón-Lagos M, Villegas VE, Rangel N et al. Tamoxifen resistance: emerging molecular targets. Int J Mol Sci 2016; 17 (8): 1357. doi: 10.3390/ijms17081357.
3. Dragomir MP, Knutsen E, Calin GA. SnapShot: unconventional miRNA functions. Cell 2018; 174 (4): 1038–e1038. doi: 10.1016/j.cell.2018.07.040.
4. Fasoulakis Z, Daskalakis G, Diakosavvas M et al. MicroRNAs determining carcinogenesis by regulating oncogenes and tumor suppressor genes during cell cycle. Microrna 2020; 9 (2): 82–92. doi: 10.2174/2211536608666190919161849.
5. Thind A, Wilson C. Exosomal miRNAs as cancer biomarkers and therapeutic targets. J Extracell Vesicles 2016; 5 (1): 3129. doi: 10.3402/jev.v5.31292.
6. Yang F, Ning Z, Ma L et al. Exosomal miRNAs and miRNA dysregulation in cancer-associated fibroblasts. Mol Cancer 2017; 16 (1): 148. doi: 10.1186/s12943-017-07 18-4.
7. Jansson MD, Lund AH. MicroRNA and cancer. Mol Oncol 2012; 6 (6): 590–610. doi: 10.1016/j.molonc.2012.09. 006.
8. Moheghi N, Afshari JT, Brook A. The cytotoxic effect of zingiber afficinale in breast cancer (MCF7) cell line. Horizon Med Sci 2011; 17 (3): 28–34.
9. Paduch R. The role of lymphangiogenesis and angiogenesis in tumor metastasis. Cell Oncol 2016; 39 (5): 397–410. doi: 10.1007/s13402-016-0281-9.
10. Jia Y, Chen Y, Wang Q et al. Exosome: emerging biomarker in breast cancer. Oncotarget 2017; 8 (25): 41717–41733. doi: 10.18632/oncotarget.16684.
11. Sun Z, Shi K, Yang S et al. Effect of exosomal miRNA on cancer biology and clinical applications. Mol Cancer 2018; 17 (1): 147. doi: 10.1186/s12943-018-0897-7.
12. Takahashi R-U, Miyazaki H, Ochiya T. The roles of microRNAs in breast cancer. Cancers 2015; 7 (2): 598–616. doi: 10.3390/cancers7020598.
13. Hannafon BN, Carpenter KJ, Berry WL et al. Exosome-mediated microRNA signaling from breast cancer cells is altered by the anti-angiogenesis agent docosahexaenoic acid (DHA). Mol Cancer 2015; 14 (1): 133. doi: 10.1186/s12943-015-0400-7.
14. Jiang M, Huang O, Zhang X et al. Curcumin induces cell death and restores tamoxifen sensitivity in the antiestrogen-resistant breast cancer cell lines MCF-7/LCC2 and MCF-7/LCC9. Molecules 2013; 18 (1): 701–720. doi: 10.3390/molecules18010701.
15. Wang P, Luo Y, Duan H et al. MicroRNA 329 suppresses angiogenesis by targeting CD146. Mol Cell Biol 2013; 33 (18): 3689–3699. doi: 10.1128/MCB.00343-13.
16. Wang L, Song G, Zheng Y et al. MiR-573 is a negative regulator in the pathogenesis of rheumatoid arthritis. Cell Mol Immunol 2016; 13 (6): 839–849. doi: 10.1038/cmi.2015.63.
17. Danza K, De Summa S, Pinto R et al. MiR-578 and miR-573 as potential players in BRCA-related breast cancer angiogenesis. Oncotarget 2015; 6 (1): 471–483. doi: 10.18632/oncotarget.2509.
Štítky
Detská onkológia Chirurgia všeobecná OnkológiaČlánok vyšiel v časopise
Klinická onkologie

2023 Číslo 6
- Brno opět přivítá onkology a nelékařské zdravotnické pracovníky
- I „pouhé“ doporučení znamená velkou pomoc. Nasměrujte své pacienty pod křídla Dobrých andělů
- Realita liečby bolesti v paliatívnej starostlivosti v Nemecku
- MUDr. Lenka Klimešová: Multiodborová vizita je kľúč k efektívnejšej perioperačnej liečbe chronickej bolesti
- Fixní kombinace tramadol/paracetamol je doporučenou volbou v léčbě chronické bolesti v ordinaci praktického lékaře
Najčítanejšie v tomto čísle
- Klasifikace zárodečných variant identifikovaných při genetickém vyšetření nádorové predispozice – konsenzus konzorcia CZECANCA
- The endoplasmic reticulum and its signaling pathways – a novel target for multiple myeloma treatment
- Nová doporučení ESMO pro klinickou praxi u metastatického kolorektálního karcinomu – komentář ke změnám v systémové léčbě
- Paliativní radioterapie pokročilého karcinomu kůže ušního boltce