
Porovnání invazivních a neinvazivních izolátů Neisseria meningitidis metodou sekvenace celého genomu, Česká republika, 2005–2021
Comparison of invasive and non-invasive isolates of Neisseria meningitidis by whole genome sequencing, Czech Republic, 2005–2021
Aim: Whole genome sequencing (WGS) analysis of candidate virulence genes of epidemiologically and/or clinically related invasive and non-invasive isolates of Neisseria meningitidis from 2005–2021.
Material and Methods: Seventy-nine isolates were selected for analysis from three different categories: cases of invasive meningococcal disease (IMD) and their healthy contacts, different clinical specimens from the same IMD case, and different clinical specimens from the same IMD case and their healthy contacts. WGS was used to analyse sequence variability in candidate N. meningitidis virulence factor genes, with more than 250 loci studied.
Results: The frequency of sequence changes in the candidate N. meningitidis virulence factor genes of invasive and non-invasive isolates varied widely. The highest level of variability was observed in the pilus genes, especially pilE and pglA. Our study detected variability in the opacity protein A (opaA) gene in more than half of the isolates analysed, with the frequency of opaA gene changes reaching almost 70% in MenC isolates. Higher frequency of changes were also observed in the genes for capsule production, especially in those of the D+D’ capsular region.
Conclusions: The results obtained support the hypothesis that serogroup-specific genetic mechanisms are also involved in the pathogenicity of N. meningitidis. These data add to the body of knowledge necessary for the development of new effective IMD vaccines.
Keywords:
Neisseria meningitidis – whole genome sequencing –virulence factors – capsular genes – pilE – pglA – opaA
Autori:
M. Honskus 1,2
![]() ; P. Křížová 1
; P. Křížová 1
![]() ; Z. Okonji 1
; Z. Okonji 1
![]() ; M. Musílek 1
; M. Musílek 1
![]() ; Kozáková
; Kozáková
Pôsobisko autorov:
Národní referenční laboratoř pro meningokokové nákazy, Centrum epidemiologie a mikrobiologie, Státní zdravotní ústav, Praha
1; 3. lékařská fakulta Univerzity Karlovy, Praha
2
Vyšlo v časopise:
Epidemiol. Mikrobiol. Imunol. 72, 2023, č. 2, s. 86-92
Kategória:
Původní práce
Súhrn
Cíl: Analýza potenciálních genů virulence metodou sekvenace celého genomu (WGS) u invazivních a neinvazivních izolátů Neisseria meningitidis příbuzných epidemiologicky a/nebo klinicky z období 2005–2021.
Materiál a metody: Pro analýzu bylo vybráno 79 izolátů ze tří různých kategorií: z případů invazivního meningokokového onemocnění (IMO) a od jejich zdravých kontaktů; z různého klinického materiálu téhož případu IMO; z různého klinického materiálu téhož případu IMO a od jejich zdravých kontaktů. Metodou WGS byly analyzovány sekvenční změny v genech potenciálních faktorů
virulence N. meningitidis, jednalo se o více než 250 lokusů.
Výsledky: Četnost sekvenčních změn v potenciálních genech virulence N. meningitidis byla u studovaných invazivních a neinvazivních izolátů značně variabilní. Nejvyšší míra genetické variability byla pozorována u genů pilu, především pilE a pglA. V naší studii byly detekovány změny v genu opacitního proteinu opaA u více než poloviny studovaných izolátů, u MenC izolátů dosahovala četnost změn genu opaA téměř 70 %. Zvýšená četnost změn byla pozorována i v genech, které jsou zodpovědné za produkci kapsule, především genech kapsulárního regionu D+D’.
Závěry: Získané výsledky podporují hypotézu, že v patogenitě N. meningitidis se uplatňují i genetické mechanismy, které jsou séroskupinově specifické. Tyto výsledky přispívají k rozšíření vědeckého poznání, které je nezbytné pro vývoj nových účinných vakcín proti IMO.
Klíčová slova:
Neisseria meningitidis – sekvenace celého genomu – faktory virulence – kapsulární geny – pilE – pglA – opaA
ÚVOD
Bakterie Neisseria meningitidis, která je výlučně lidským patogenem, kolonizuje horní cesty dýchací a většinou nezpůsobuje závažná onemocnění. Zatímco nosičství N. meningitidis je relativně běžné (zjišťováno až u 10 % zdravé populace), invazivní meningokokové onemocnění (IMO) je vzácné. Závažnost IMO tkví ve vysoké smrtnosti (až 20 %) a velkém procentu těžkých celoživotních následků u přeživších. Nebezpečnost IMO také spočívá ve skutečnosti, že se během několika hodin z plného zdraví může rozvinout život ohrožující onemocnění. Většina IMO se objevuje jako sporadické případy, bez prokázaného kontaktu s jiným IMO [1].
Faktory, které ovlivňují rozvoj bezpříznakového nosičství N. meningitidis či rozvoj závažného, život ohrožujícího IMO, jsou popsány jak na straně hostitele (například věk, poruchy imunity, socioekonomické faktory, kuřáctví, psychické či fyzické oslabení), tak na straně meningokoka (například velikost infekční dávky, příslušnost k hypervirulentnímu klonálnímu komplexu, přítomnost faktorů virulence). Ačkoli byla popsána řada mechanismů důležitých pro virulenci N. meningitidis, stále ještě nejsou definovány faktory, které by jasně odlišily vysoce patogenní izoláty působící IMO od méně patogenních [2, 3].
Izoláty N. meningitidis z IMO jsou v naprosté většině zařazeny do některé ze šesti sérologických skupin (A, B, C, X, Y, W) a velmi často patří k hypervirulentním klonálním komplexům, které působí celosvětově většinu IMO. Meningokoky izolované od zdravých nosičů jsou geneticky vysoce heterogenní, až u 30 % z nich nelze určit séroskupinu a většinou patří k nevirulentním klonálním komplexům. Přítomnost polysacharidové kapsule zvyšuje šanci k vyvolání IMO, zatímco meningokok bez kapsule pravděpodobněji způsobí pouze kolonizaci sliznic hostitele. V rámci rodu Neisseria můžeme expresi kapsulárního polysacharidu nalézt výhradně u N. meningitidis a je vysoce pravděpodobné, že získání genetického materiálu potřebného pro produkci kapsule bylo důležitým krokem v evoluci meningokoka [4]. Geny zodpovědné za syntézu a transport kapsule jsou umístěny v kapsulárních regionech: A, B, C a D+D‘ [5]. Nové studie prokázaly, že izoláty N. meningitidis z IMO, které byly dříve neodlišitelné od nosičských izolátů pomocí klasických laboratorních typizačních metod, jsou nyní rozlišitelné díky nové metodě sekvenace celého genomu (Whole Genome Sequencing, WGS) [6]. Studium metodou WGS poskytuje důkazy o tom, že virulence N. meningitidis je polygenní vlastností a přispívá do ní různou mírou mnoho rozdílných genů [7].
Předkládáme výsledky analýzy potenciálních genů virulence metodou WGS u 79 invazivních a neinvazivních izolátů N. meningitidis příbuzných epidemiologicky a/nebo klinicky z období 2005–2021.
MATERIÁL A METODY
Izoláty N. meningitidis
Výběr izolátů byl proveden na základě epidemiologických a klinických informací o izolátech N. meningitidis v databázi Národní referenční laboratoře (NRL) pro meningokokové nákazy, která obsahuje i data získaná klasickou sekvenací [8, 9, 10]. Celosvětově je k dispozici minimum sbírek izolátů N. meningitidis, které disponují detailními epidemiologickými, klinickými a mikrobiologickými daty – česká sbírka je z tohoto hlediska unikátní.
Pro studium potenciálních faktorů virulence izolátů N. meningitidis metodou WGS byly ze sbírky NRL vybrány izoláty příbuzné epidemiologicky a/nebo klinicky z období 2005–2021, které byly zařazeny do tří kategorií:
A) z případů IMO a od jejich zdravých kontaktů (nosičů N. meningitidis)
– 11 souborů, 28 izolátů
B) z různého klinického materiálu téhož případu IMO (likvor, krev, horní cesty dýchací)
– 14 souborů, 30 izolátů
C) z různého klinického materiálu téhož případu IMO a od jejich zdravých kontaktů
– 7 souborů, 21 izolátů
Celkem: 32 srovnávaných souborů, 79 izolátů N. meningitidis.
Do těchto souborů invazivních a neinvazivních izolátů příbuzných epidemiologicky a/nebo klinicky byly vždy vybrány pouze izoláty vzájemně neodlišitelné klasickými typizačními metodami (séroskupina, sekvenační typ, klonální komplex).
Celogenomová sekvenace N. meningitidis
Příslušné bakteriální kultury N. meningitidis, které jsou uchovávány ve sbírce NRL pro meningokokové nákazy, byly vyočkovány na čokoládový Mueller-Hinton agar a kultivovány 18–24 hodin ve 37 °C v 5% CO2 atmosféře. Správnost identifikace N. meningitidis byla ověřena pomocí kitu API NH (BIOMÉRIEUX). Séroskupiny byly určeny klasickými sérologickými metodami (Pastorex Meningitis Bio-RAD, antiséra N. meningitidis ITEST, Bio-RAD) a ověřeny metodou RT-PCR. K izolaci DNA byla použita souprava QIAamp DNA Mini Kit (QIAGEN) a postup izolace probíhal podle pokynů výrobce. DNA byla následně odeslána k celogenomové sekvenaci na externí pracoviště EMBL (European Molecular Biology Laboratory, Heidelberg, Německo). Sekvenace proběhla na platformě Illumina MiSeq s využitím referenčního genomu N. meningitidis kmene MC58 a výsledkem byly překrývající se sekvence o délce přibližně 300 bp. K sestavení výsledných genomů z primárních celogenomových dat byl na našem pracovišti použit software Velvet de novo Assembler [11]. Genomy izolátů byly poté registrovány v databázi PubMLST, která využívá platformu BIGSdb (Bacterial Isolate Genome Sequence Database) [12, 13].
Výběr potenciálních genů virulence N. meningitidis
Výběr studovaných genů byl učiněn na základě rešerše literatury a dle informací v databázi PubMLST. Jednalo se o více než 250 lokusů, z nichž většina je v databázi PubMLST ve vyhledávacích schématech, která sdružují lokusy s ohledem na funkci jejich proteinových produktů. Hodnoceno bylo 7 lokusů u 5 antigenních genů (fetA, nadA, nhba, porA, porB), 11 lokusů u 8 genů antibiotické rezistence (folP, gyrA, mtrR, parC, parE, penA, ponA, rpoB), 3–7 lokusů kapsulárního regionu A, které jsou séroskupinově specifické, dále lokusy zbylých kapsulárních regionů – region B (ctrE, ctrF), region C (ctrA, ctrB, ctrC, ctrD) a region D+D‘ (galE, galE2, rfbA, rfbB, rfbC, rfbC2). Testována byla přítomnost a změny gonokokových genetických ostrovů (73 lokusů) a plazmidů – konjugativních (50 lokusů), kryptických (9 lokusů) a β-laktamázových (8 lokusů). Dále bylo hodnoceno 11 genů glykolýzy (eno, fba, fbp, gapA, gapA2, gpm, pgi1, pgi2, pgk, pgm, pykA), 7 genů pro proteiny spojené s akvizicí železa (hmbR, hpuA, hpuB, lbpA, lbpB, tbpA, tbpB), 19 genů zapojených do metabolismu lipooligosacharidů (LOS) – α-chain transferáz (lgtA, lgtB, lgtC, lgtD, lgtE, lgtF, lst), inner core transferáz (lgtG, lot, lpt3, lpt6, rfaC, rfaF, rfaK) a geny proteinů transportu/exportu LOS (lptC, msbA, ostA, rlpB, yhbG). Výčet hodnocených lokusů uzavírá 27 genů pilu (pilA, pilB, pilC1, pilC2, pilD, pilE, pilF, pilG, pilH, pilI, pilJ, pilK, pilM, pilN, pilO, pilP, pilQ, pilS, pilT1, pilT2, pilU, pilV, pilW, pilX, pilZ, pglA, comP), 4 geny ze skupiny opa a opc (opaA, opaB, opcA, opcB), 8 genů lpx (lpxA, lpxB, lpxC, lpxD, lpxH, lpxK, lpxL, lpxL2) a gen iga2, jehož produktem je serinová peptidáza imunoglobulinu A.
VÝSLEDKY A DISKUSE
Pro analýzu genů potenciálních faktorů virulence bylo vybráno celkem 79 izolátů N. meningitidis s následujícím zastoupením séroskupin: séroskupina C (MenC, 34 izolátů), séroskupina B (MenB, 31 izolátů), séroskupina W (MenW, 9 izolátů), séroskupina Y (MenY, 2 izoláty) a tři izoláty nebyly přiřazeny k žádné séroskupině (non-groupable, MenNG). U dvou z těchto MenNG byla metodou WGS prokázána genoskupina X, u jednoho genoskupina B s inaktivovanou kapsulární polymerázou. Z pohledu srovnávaných příbuzných souborů se jednalo o 13 souborů izolátů MenC, 13 souborů izolátů MenB včetně jednoho příbuzného izolátu MenNG a 6 souborů izolátů ostaních séroskupin/genoskupin (MenW, MenY a MenX). Izoláty studovaného souboru náležely do 8 klonálních komplexů: cc11 (30 izolátů), cc41/44 (15 izolátů), cc32 (5 izolátů), cc865 (5 izolátů), cc22 (4 izoláty), cc35 (3 izoláty), cc269 (3 izoláty), cc174 (2 izoláty), cc750 (2 izoláty). U 10 izolátů byl zjištěn sekvenační typ (ST), který v databázi PubMLST není přiřazen do žádného z klonálních komplexů (unassigned, ccUA).
Mezi 79 studovanými invazivními a neinvazivními izoláty N. meningitidis nebyly v rámci jednotlivých příbuzných souborů detekovány žádné sekvenční změny u genů antibiotické rezistence, genů, které kódují proteiny transportu/exportu LOS, a genů kapsulárního regionu C, které se podílejí na translokaci kapsulárního polysacharidu s vysokou molekulovou hmotností na buněčný povrch (obr. 1). Žádné změny nebyly rovněž pozorovány v případě plazmidů, jejichž přítomnost byla potvrzena pouze u jednoho srovnávaného souboru izolátů (2 lokusy ze skupiny kryptických plazmidů), a gonokokových genetických ostrovů, které byly detekovány u dvou souborů (60 a 61 různých lokusů). Minimální míra sekvenčních změn byla zjištěna u antigenních genů, genů glykolýzy, genu iga2 a genů kapsulárního regionu B, které jsou zapojeny do translokace kapsulárního polysacharidu. Vyšší variabilitu vykazovaly séroskupinově specifické geny kapsulárního regionu A, které jsou nezbytné pro syntézu kapsulárního polysacharidu, geny LOS α-chain transferáz (zejména geny lgtA, lgtC a lgtD), geny lpx (především gen lpxL) a v případě genů LOS inner core transferáz byly změny zaznamenány výhradně u genu lgtG (u 8 souborů z 32). Nejvýraznější genetická variabilita byla pozorována u genů pilu (téměř 80 % studovaných souborů), genů ze skupiny opa a opc (více než 50 % souborů), genů klíčových pro akvizici železa a genů kapsulárního regionu D+D‘, jejichž produkty se účastní syntézy kapsulárního polysacharidu.
Figure. 1. Frequency of changes in candidate N. meningitidis virulence factor genes in 32 groups of invasive and
non-invasive isolates, by gene function

Vysoká genetická variabilita genů pilu byla z velké části tvořena sekvenčními změnami u genů pilE a pglA (obr. 2). Ostatní z celkových 27 genů pilu nevykazovaly žádné (pilA, pilB, pilD, pilG, pilH, pilI, pilJ, pilK, pilM, pilN, pilO, pilP, pilT1, pilT2, pilV, pilW, pilX, pilZ, comP), nebo jen minimální změny (pilC1, pilC2, pilF, pilQ, pilS, pilU). V případě rozdělení studovaných souborů příbuzných izolátů podle příslušných séroskupin lze u izolátů MenB pozorovat vyšší četnost změn u genu pilE, oproti tomu u izolátů MenC dominovaly změny genu pglA (obr. 3). U izolátů ostatních séroskupin byla četnost změn v genech pilE a pglA srovnatelná.
Figure 2. Frequency of sequence changes in 27 N. meningitidis pilus genes

Figure 3. Frequency of sequence changes in selected eight pilus genes, by serogroup

Detailnější pohled na změny u 4 genů skupiny opa a opc odhalil skutečnost, že genetická variabilita, zjištěná v rámci této skupiny genů, byla způsobena výhradně sekvenčními změnami genu pro opacitní protein opaA (obr. 4). Změny v sekvenci opaA byly zjištěny u více než poloviny studovaných souborů izolátů, oproti tomu ostatní geny skupiny opa a opc (opaB, opcA, opcB) žádnou variabilitu nevykazovaly. Rozdíl v četnosti změn ve studovaných souborech byl také pozorován po rozdělení podle séroskupin. Zatímco u MenB izolátů vykazoval gen opaA změnu u více než 50 % a u izolátů ostatních séroskupin více než u 30 % studovaných souborů, u MenC izolátů dosahovala četnost změn u genu opaA téměř 70 %.
Figure 4. Frequency of sequence changes in the opa and opc
genes, by serogroup for the opaA gene

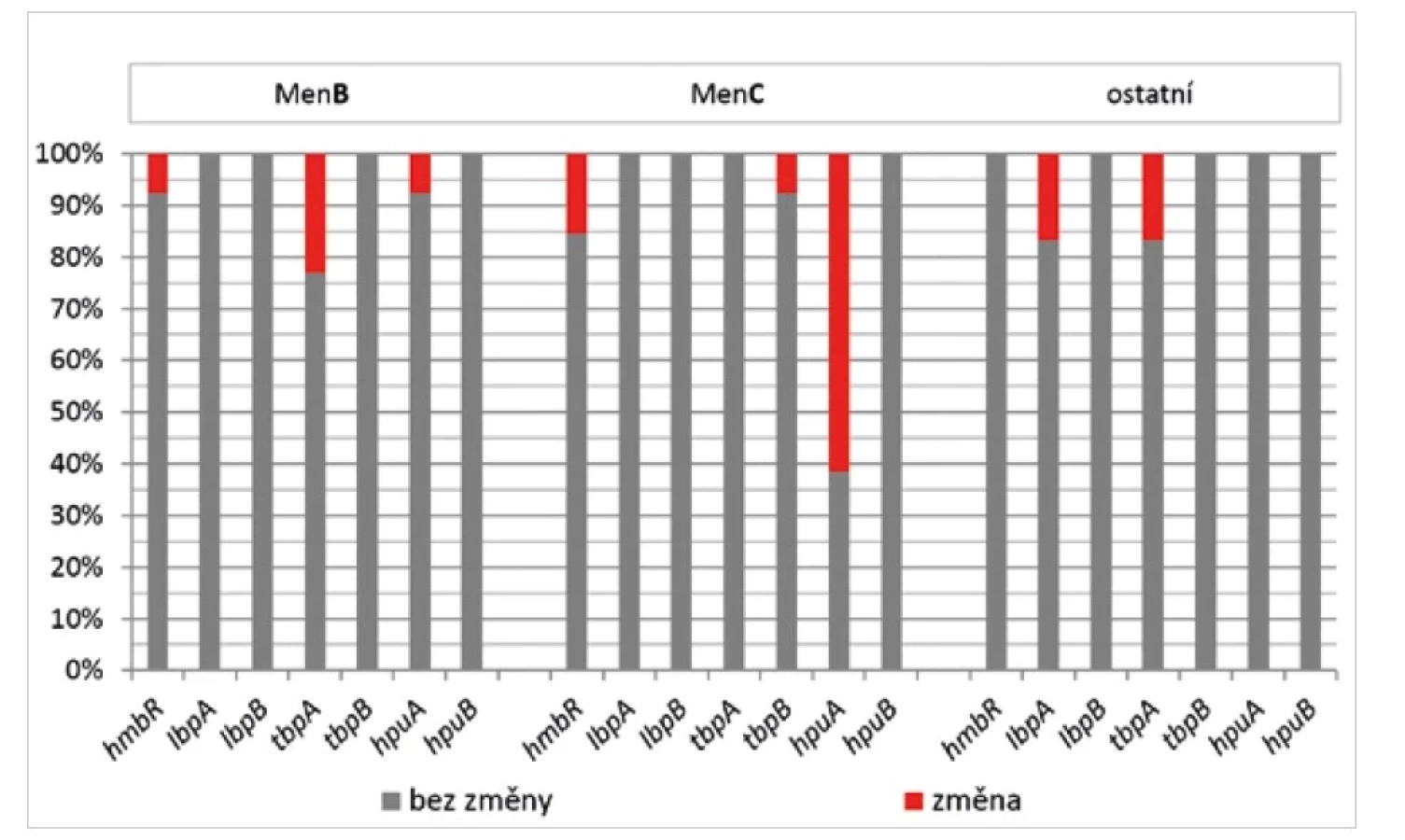
Skoro polovina studovaných souborů izolátů vykazovala změny u genů, které jsou klíčové pro akvizici železa (viz obr. 1). V tomto případě byla genetická variabilita rozložena mezi 5 různých genů, pouze u genů lbpB a hpuB nebyly v jednotlivých souborech pozorovány žádné změny (obr. 5). Při rozdělení podle příslušných séroskupin lze u izolátů MenB pozorovat mírně zvýšenou frekvenci změn v genu tbpA, zatímco u izolátů MenC dominují změny u genu hpuA, který kóduje hemoglobin-haptoglobin utilizační protein.
Figure 5. Frequency of sequence changes in the N. meningitidis iron acquisition genes, by serogroup
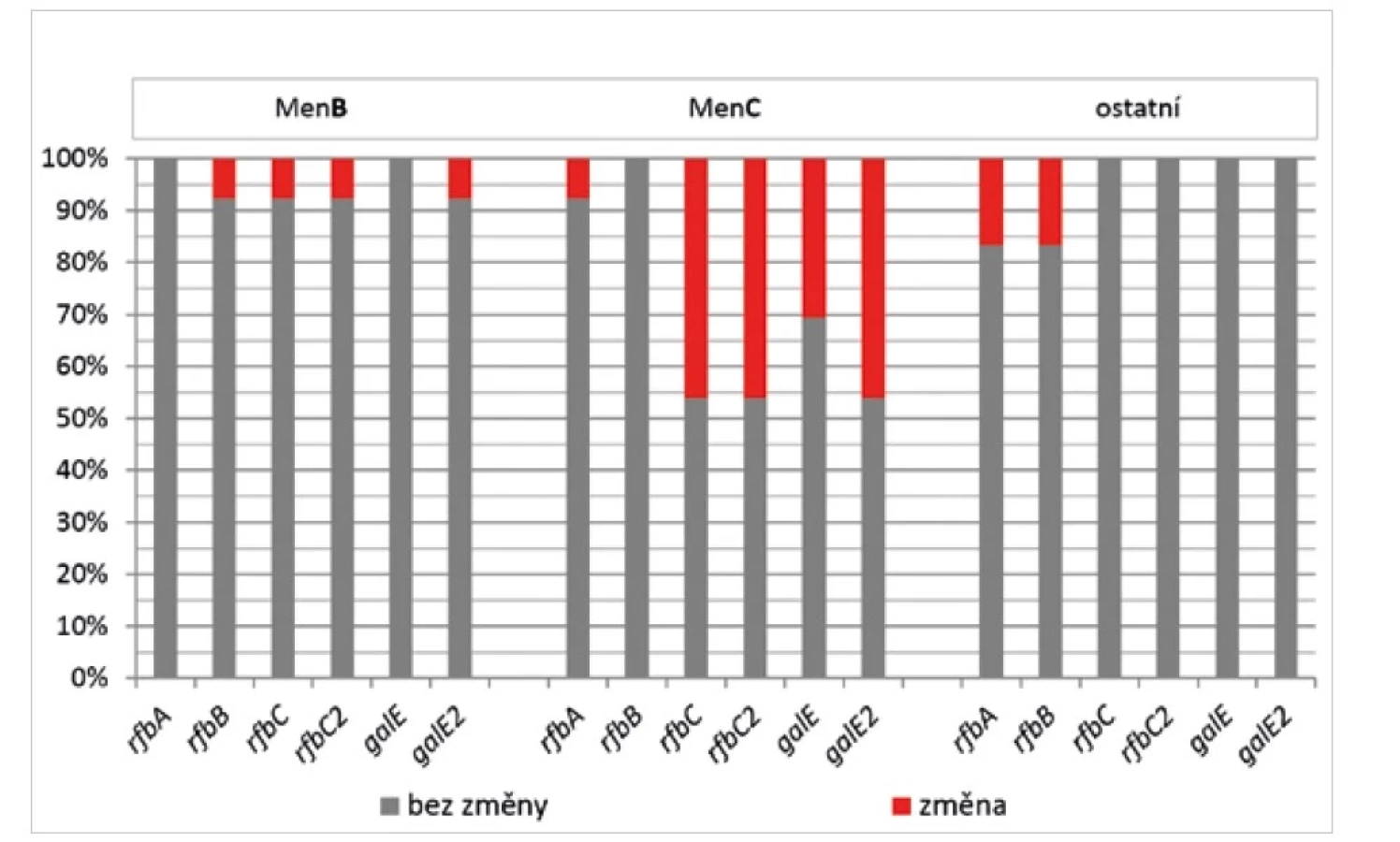
Vysoká variabilita, zejména z pohledu změn u více než jednoho genu (viz obr. 1), byla v rámci studovaných souborů invazivních a neinvazivních izolátů zaznamenána u všech 6 genů kapsulárního regionu D+D‘ (obr. 6). U izolátů MenB byla četnost změn minimální, oproti tomu u MenC izolátů byla tato četnost výrazně vyšší, zejména u genů rfbC, rfbC2, galE a galE2. U izolátů ostatních séroskupin byly naopak zjištěny změny pouze u genů rfbA a rfbB.
Figure 6. Frequency of sequence changes in six genes of the D+D’ capsular region, by serogroup
Souhrnně lze říci, že u potenciálních genů virulence N. meningitidis byla četnost sekvenčních změn v rámci studovaných souborů invazivních a neinvazivních příbuzných izolátů značně variabilní. Zatímco u mnoha skupin studovaných genů byly zjištěny minimální nebo žádné změny, u několika skupin genů byla četnost změn mezi invazivními a neinvazivními izoláty vysoká. Nejvyšší míra genetické variability byla pozorována u genů pilu, který se řadí mezi hlavní adhezivní molekuly a představuje významný faktor virulence N. meningitidis [14]. Dominantní podíl na této variabilitě měly geny pilE a pglA. Vysoká míra sekvenční variability genu pilE, který kóduje hlavní subjednotku pilu, je dobře popsána [15, 16] a vychází ze schopnosti genu pilE rekombinovat s inaktivními kopiemi genu pilS [17]. Gen pglA se účastní post-translační glykosylace pilu a podléhá fázovým variacím v důsledku přítomnosti poly-G úseku ve své sekvenci [14, 17, 18]. V našem souboru byla u izolátů MenB prokázána vyšší četnost změn genu pilE, oproti tomu u izolátů MenC dominovaly změny genu pglA. U izolátů ostatních séroskupin byla četnost změn v genech pilE a pglA srovnatelná. Opacitní proteiny skupiny opa a opc jsou hlavní proteiny vnější membrány N. meningitidis a přispívají k patogenitě meningokoků svoji významnou rolí v adhezi bakterií k buňkám nosohltanu a modulaci imunitního systému díky interakci s imunokompetentními buňkami [19, 20]. Jejich význam potvrzuje i to, že v minulosti patřily tyto proteiny mezi vakcinační kandidáty [21]. V naší studii byly detekovány změny v sekvenci genu opaA u více než poloviny studovaných izolátů, v případě MenC izolátů dosahovala četnost změn genu opaA téměř 70 %. Možnou příčinou tak vysoké genetické variability může být přítomnost transformačního hot-spotu v blízkosti genu opaA [22, 23]. Izoláty klinicky nejvýznamnějších séroskupin B, C, W a Y produkují na svůj povrch kapsulární polysacharid obohacený kyselinou sialovou, který hraje klíčovou roli v patogenitě N. meningitidis [24]. Zvýšená četnost změn v genech, které jsou zodpovědné za produkci kapsule, byla pozorována i v souborech námi studovaných izolátů, což odpovídá skutečnosti, že N. meningitidis disponuje řadou genetických mechanismů k regulaci produkce kapsule [25]. Zjištěná genetická variabilita byla výrazně vyšší u genů, které se účastní syntézy kapsulárního polysacharidu (kapsulární region A a zejména region D+D‘), než u genů zapojených do translokace polysacharidu na buněčný povrch (kapsulární regiony B a C). Výsledky zjištěné v této studii zároveň podporují hypotézu, že v patogenitě N. meningitidis se uplatňují i genetické mechanismy, které jsou séroskupinově specifické [26]. V naší studii se jednalo především změny v genech pilu a genech kapsulárního regionu D+D‘.
ZÁVĚRY
Četnost sekvenčních změn v potenciálních genech virulence N. meningitidis byla u studovaných invazivních a neinvazivních izolátů značně variabilní. Nejvyšší míra genetické variability byla pozorována u genů pilu, především pilE a pglA. V naší studii byly detekovány změny v genu opacitního proteinu opaA u více než poloviny studovaných izolátů, u MenC izolátů dosahovala četnost změn genu opaA téměř 70 %. Zvýšená četnost změn byla pozorována i v genech, které jsou zodpovědné za produkci kapsule, především genech kapsulárního regionu D+D‘. Získané výsledky podporují hypotézu, že v patogenitě N. meningitidis se uplatňují i genetické mechanismy, které jsou séroskupinově specifické. Tyto výsledky přispívají k rozšíření vědeckého poznání, které je nezbytné pro vývoj nových účinných vakcín proti IMO.
Podpora projektu
Podpořeno z programového projektu Ministerstva zdravotnictví ČR s reg. č. NV19-09-00319.
Do redakce došlo dne 20. 12. 2022.
Adresa pro korespondenci:
MUDr. Pavla Křížová, CSc.
SZÚ Praha
Šrobárova 49/48
100 42 Praha 10
e-mail: pavla.krizova@szu.cz
Zdroje
1. Caugant DA, Tzanakaki G, Kriz P. Lessons from meningococcal carriage studies. FEMS Microbiol Rev. 2007;31(1):52–63. Doi: 10.1111/j.1574-6976.2006.00052.x. PMID: 17233635.
2. Tan A, Hill DM, Harrison OB, et al. Distribution of the type III DNA methyltransferases modA, modB and modD among Neisseria meningitidis genotypes: implications for gene regulation and virulence. Sci Rep. 2016;6 : 21015. Doi: 10.1038/srep21015. PMID: 26867950.
3. Bårnes GK, Brynildsrud OB, Børud B, et al. Whole genome sequencing reveals within host genetic changes in paired meningococcal carriage isolates from Ethiopia. BMC Genomics, 2017;18 : 407. Doi: 10.1186/s12864-017-3806-3.
4. Claus H, Maiden MCJ, Maag R, et al. Many carried meningococci lack the genes required for capsule synthesis and transport. Microbiology (Reading). 2002;148(6):1813–1819. Doi: 10.1099/00221287-148-6-1813. PMID: 12055301.
5. Jones CH, Mohamed N, Rojas R, et al. Comparison of Phenotypic and Genotypic Approaches to Capsule Typing of Neisseria meningitidis by Use of Invasive and Carriage Isolate Collections. J. Clin. Microbiol., 2016;54(1):25–34. Doi: 10.1128/JCM.01447-15. PMID: 26311858.
6. Ren X, Eccles DA, Greig GA, et al. Genomic, Transcriptomic, and Phenotypic Analyses of Neisseria meningitidis Isolates from Disease Patients and Their Household Contacts. mSystems, 2017;2(6):e00127–17. Doi: 10.1128/mSystems.00127-17. PMID: 29152586.
7. Joseph B, Schneiker-Bekel S, Schramm-Glück A, et al. Comparative genome biology of a serogroup B carriage and disease strain supports a polygenic nature of meningococcal virulence. J Bacteriol., 2010;192(20):5363–77. Doi: 10.1128/JB.00883-10. PMID: 20709895.
8. Maiden MC, Bygraves JA, Feil E, et al. Multilocus sequence typing: a portable approach to the identification of clones within populations of pathogenic microorganisms. Proc. Natl. Acad. Sci. U S A, 1998;95(6):3140–3145. Doi: 10.1073/pnas.95.6.3140. PMID: 9501229.
9. Brehony C, Jolley KA, Maiden MC. Multilocus sequence typing for global surveillance of meningococcal disease. FEMS Microbiol. Rev., 2007;31(1):15–26. Doi: 10.1111/j.1574-6976.2006.00056.x. PMID: 17168997.
10. Jolley KA, Maiden MC. Using multilocus sequence typing to study bacterial variation: prospects in the genomic era. Future Microbiol., 2014;9(5):623–630. Doi: 10.2217/fmb.14.24. PMID: 24957089.
11. Zerbino DR. Using the Velvet de novo assembler for shortread sequencing technologies. Curr. Protoc. Bioinformatics, 2010;11:Unit 11.5. Doi: 10.1002/0471250953.bi1105s31. PMID: 20836074.
12. Jolley KA, Maiden MC. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinformatics, 2010;11 : 595. Doi: 10.1186/1471-2105-11-595. PMID: 21143983.
13. Jolley KA, Bray JE, Maiden MCJ. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res., 2018;3 : 124. Doi: 10.12688/wellcomeopenres.14826.1. PMID: 30345391.
14. Jennings MP, Virji M, Evans D, et al. Identification of a novel gene involved in pilin glycosylation in Neisseria meningitidis. Mol. Microbiol., 1998;29(4):975–984. Doi: 10.1046/j.1365-2958.1998.00962.x. PMID: 9767566.
15. Rytkönen A, Albiger B, Hansson-Palo P, et al. Neisseria meningitidis undergoes PilC phase variation and PilE sequence variation during invasive disease. J. Infect. Dis., 2004;189(3):402–409. Doi: 10.1086/381271. PMID: 14745697.
16. Sun X, Zhou H, Xu L, et al. Prevalence and genetic diversity of two adhesion-related genes, pilE and nadA, in Neisseria meningitidis in China. Epidemiol Infect. 2013;141(10):2163–2172. Doi: 10.1017/S0950268812002944. PMID: 23290624.
17. Carbonnelle E, Hill DJ, Morand P, et al. Meningococcal interactions with the host. Vaccine, 2009;27(Suppl 2):B78–89. Doi: 10.1016/j.vaccine.2009.04.069. PMID: 19481311.
18. Power PM, Roddam LF, Rutter K, et al. Genetic characterization of pilin glycosylation and phase variation in Neisseria meningitidis. Mol Microbiol. 2003;49(3):833–847. Doi: 10.1046/j.1365-2958.2003.03602.x. PMID: 12864863.
19. Sadarangani M, Pollard AJ, Gray-Owen SD. Opa proteins and CEACAMs: pathways of immune engagement for pathogenic Neisseria. FEMS Microbiol. Rev., 2011;35(3):498–514. Doi: 10.1111/j.1574-6976.2010.00260.x. PMID: 21204865.
20. Gasparini R, Amicizia D, Lai PL, et al. Neisseria meningitidis, pathogenetic mechanisms to overcome the human immune defences. J. Prev. Med. Hyg., 2012;53(2):50–55. PMID: 23240160.
21. Sadarangani M, Hoe JC, Callaghan MJ, et al. Construction of Opa-positive and Opa-negative strains of Neisseria meningitidis to evaluate a novel meningococcal vaccine. PLoS One, 2012;7(12):e51045. Doi: 10.1371/journal.pone.0051045. PMID: 23251421.
22. Claus H, Frosch M, Vogel U. Identification of a hotspot for transformation of Neisseria meningitidis by shuttle mutagenesis using signature-tagged transposons. Mol. Gen. Genet., 1998;259(4):363–371. Doi: 10.1007/s004380050823. PMID: 9790590.
23. Linz B, Schenker M, Zhu P, et al. Frequent interspecific genetic exchange between commensal Neisseriae and Neisseria meningitidis. Mol Microbiol. 2000;36(5):1049–1058. Doi: 10.1046/j.1365-2958.2000.01932.x. PMID: 10844690.
24. Tzeng YL, Swartley JS, Miller YK, et al. Transcriptional regulation of divergent capsule biosynthesis and transport operon promoters in serogroup B Neisseria meningitidis. Infect Immun., 2001;69(4):2502–2511. Doi: 10.1128/IAI.69.4.2502-2511.2001. PMID: 11254613.
25. Tzeng YL, Thomas J, Stephens DS. Regulation of capsule in Neisseria meningitidis. Crit. Rev. Microbiol., 2016;42(5):759–772. Doi: 10.3109/1040841X.2015.1022507. PMID: 26089023.
26. Talà A, Cogli L, De Stefano M, et al. Serogroup-specific interaction of Neisseria meningitidis capsular polysaccharide with host cell microtubules and effects on tubulin polymerization. Infect Immun., 2014;82(1):265–274. Doi: 10.1128/IAI.00501-13. PMID: 24166951.
Štítky
Hygiena a epidemiológia Infekčné lekárstvo MikrobiológiaČlánok vyšiel v časopise
Epidemiologie, mikrobiologie, imunologie

2023 Číslo 2
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Očkování proti virové hemoragické horečce Ebola experimentální vakcínou rVSVDG-ZEBOV-GP
- Koronavirus hýbe světem: Víte jak se chránit a jak postupovat v případě podezření?
Najčítanejšie v tomto čísle
- Slovo o mikrobiomu: úvaha nad historií, současným stavem a pojmoslovím vzkvétajícího oboru
- Vybrané aspekty úmrtnosti v Česku a na Slovensku v pandemickém roce 2020
- The influence of meteorological factors on the risk of tick-borne encephalitis infection
- Sekundární prevence karcinomu plic v České republice – úskalí, rizika, benefit