
Sila pôstu: metabolické, hormonálne a psychologické účinky prerušovaného lačnenia
The power of starvation: metabolic, hormonal, and psychological effects of intermittent fasting
Available clinical evidence suggests that intermittent fasting (IF), including time-restricted eating, may improve certain parameters of metabolic health in selected patients (body weight, glycemia/insulin sensitivity, and some cardiometabolic risk markers). The effect is often also related to energy deficit, diet quality, and circadian timing of meals. Data on the reduction of “subclinical” inflammation are heterogeneous (changes in some biomarkers are observed, but not consistently across studies). IF may induce a physiological “metabolic switch” toward increased fat oxidation/ketosis; however, preservation of muscle mass is not automatic and requires adequate protein intake and resistance training. Mechanistically, IF activates pathways associated with stress adaptation and autophagy, but direct clinical evidence of tissue autophagy in humans is limited. Evidence regarding neuroprotection, mood, and sleep is promising but currently inconclusive and highly individual. IF is not a universal solution; the choice of protocol, goals, and safety precautions should be individualized, with increased caution and often medical supervision in high-risk groups (especially in patients on antidiabetic therapy with risk of hypoglycemia, during pregnancy, in individuals with eating disorders, and in frail underweight patients).
Keywords:
Starvation – autophagy – fasting – intermittent fasting – metabolic effects
Authors:
Emil Martinka
Authors‘ workplace:
Diabetologické oddelenie NEDÚ n. o., Ľubochňa
Published in:
Forum Diab 2026; 15(1): 26-35
Category:
Topic
Overview
Dostupné klinické dôkazy naznačujú, že prerušované lačnenie (IF – Intermittent Fasting) vrátane časovo obmedzeného jedla (time-restricted eating) môže u vybraných pacientov zlepšiť niektoré parametre metabolického zdravia (telesnú hmotnosť, glykémiu/inzulínovú senzitivitu a časť kardiometabolických rizikových markerov), pričom účinok často súvisí aj s energetickým deficitom, kvalitou stravy a cirkadiánnym načasovaním jedál. Dáta o znížení „subklinického“ zápalu sú heterogénne (zmeny niektorých biomarkerov sa pozorujú, no nie konzistentne naprieč štúdiami). IF môže navodiť fyziologický metabolický „switch“ smerom k vyššej oxidácii tukov/ketóze, avšak zachovanie svalovej hmoty nie je automatické a vyžaduje adekvátny príjem proteínu a silový tréning. Mechanistické vysvetlenie je, že IF aktivuje dráhy spojené so stresovou adaptáciou a autofágiou, no priame klinické dôkazy o tkanivovej autofágii u ľudí sú obmedzené. Údaje o neuroprotekcii, nálade a spánku sú sľubné, ale zatiaľ nejednoznačné a výrazne individuálne. IF nie je univerzálne riešenie; výber protokolu, cieľov a bezpečnostných opatrení má byť individualizovaný, so zvýšenou opatrnosťou a často aj s medicínskym dohľadom u rizikových skupín (najmä pri antidiabetickej liečbe s rizikom hypoglykémie, v gravidite, pri poruchách príjmu potravy a u krehkých pacientov s telesnou podhmotnosťou).
Klíčová slova:
autofágia – prerušované lačnenie (intermittent fasting – IF) – lačnenie – metabolické účinky – pôst
Úvod
Stravovanie sa v moderných spoločnostiach je charakteristické častým príjmom potravy počas dňa, často presahujúcim 14–16 hodín denne. Tento stravovací model u mnohých osôb (najmä pri nadbytku energie, inzulínovej rezistencii a večernom príjme) vedie ku chronickému postprandiálnemu metabolickému stavu, sprevádzanému dlhodobo zvýšenou hladinou glukózy, zvýšenou sekréciou inzulínu, potlačením lipolýzy a obmedzeným využívaním vlastných tukových zásob. Chronická hyperinzulinémia je súčasťou komplexného mechanizmu, ktorý podporuje lipogenézu, ektopické ukladanie tuku, rozvoj inzulínovej rezistencie, čím významne prispieva k progresii obezity, metabolického syndrómu a ním indukovaných ochorení vrátane diabetes mellitus 2. typu (DM2T) [1–3].
Intermitentné lačnenie
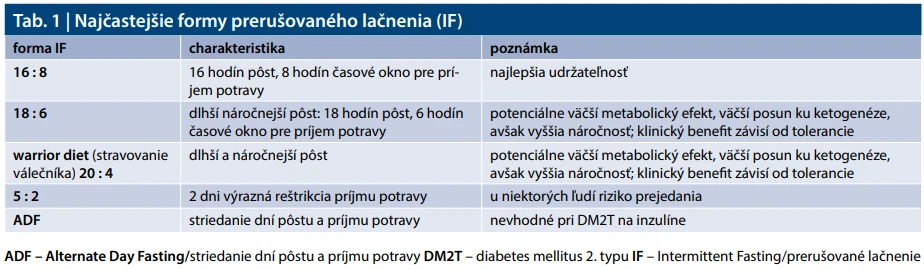
Intermitentné dobrovoľné (cielené) lačnenie (IF – Intermittent Fasting), resp. prerušované lačnenie/pôst, predstavuje návrat k fyziologickému striedaniu období príjmu potravy a lačnenia, ktoré bolo pre človeka evolučne prirodzené. Na rozdiel od kontinuálnej kalorickej reštrikcie IF vytvára opakované obdobia nízkeho príjmu energie a nízkej inzulinémie, čím môže aktivovať pôstom podmienené adaptačné mechanizmy ako sú zvýšená lipolýza, ketogenéza a signalizácia AMPK (Adenosine Monophosphate-activated Protein Kinase) – mTOR (mechanictic Target Of Rapamycin), aj bez potreby nepretržitého každodenného kalorického obmedzenia [4,5]. Klinické metabolické benefity IF sú však často aspoň čiastočne sprostredkované celkovým energetickým deficitom a zmenou telesnej hmotnosti. Tab. 1 sumarizuje najčastejšie formy IF s dĺžkou lačnenia a časovým oknom pre príjem potravy.
Metabolické fázy lačnenia (fyziologický kontext)
Počas lačnenia metabolizmus prechádza viacerými fázami, počas ktorých sa energetický metabolizmus postupne a do veľkej miery posúva od využívania energie z glukózy prijatej v potrave k energii získavanej z mastných kyselín a ketolátok vlastných tukových zásob. Toto presmerovanie sa označuje ako metabolický „switch“, ktorý nastáva pri pôste a ku ktorému dochádza po vyčerpaní zásob glykogénu. Schopnosť organizmu prepínať medzi jednotlivými zdrojmi energie sa označuje ako metabolická flexibilita a je vlastnosťou metabolického zdravia. Ak má organizmus dobrú metabolickú flexibilitu, metabolický switch nastane ľahšie a rýchlejšie, naopak, ak je prítomná inzulínová rezistencia, resp. metabolický syndróm, prepnutie je pomalšie a menej efektívne.
Metabolický switch je sprevádzaný poklesom inzulinémie a zvýšenou oxidáciou tukov. Pri opakovanom a dlhodobejšom praktizovaní IF sa metabolická flexibilita zlepšuje, telo sa „učí“ efektívnejšie využívať tuky. Môže viesť k zlepšeniu inzulínovej senzitivity, k poklesu hladín triglyceridov, viscerálneho tuku a HbA1c. Aktivácia autofágie a potenciálne neuroprotektívne účinky sú mechanisticky dobre podložené, avšak u ľudí zostáva úroveň klinických dôkazov zatiaľ limitovaná. Komplexný prehľad zmien počas lačnenia sumarizuje tab. 2.
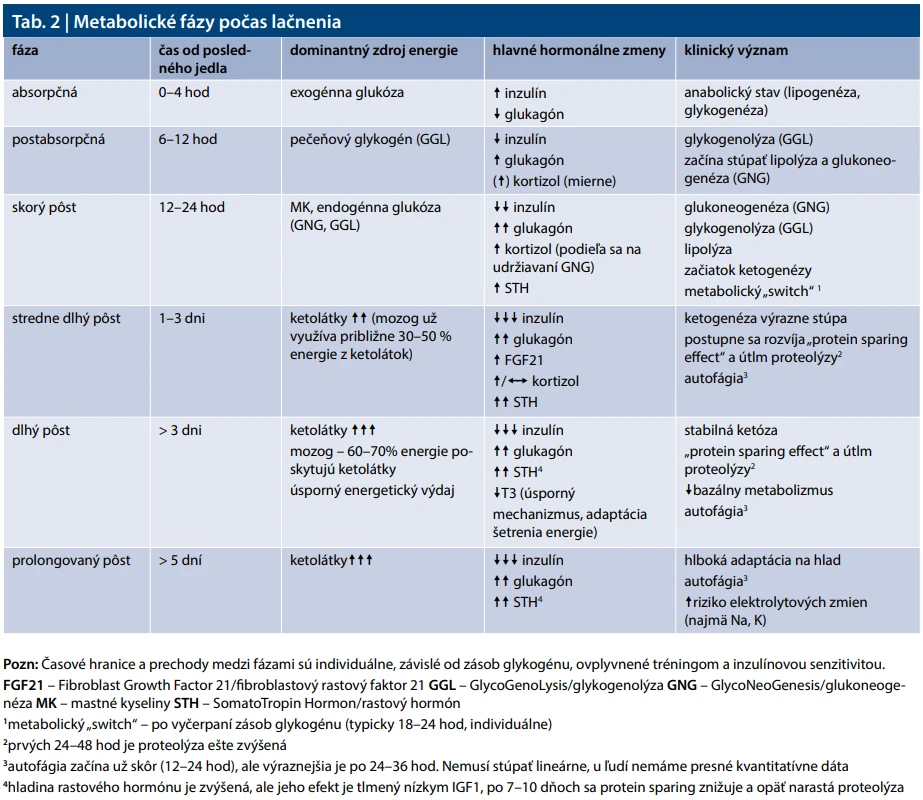
Absorpčná fáza (< 4 hodiny po jedle)
Po príjme potravy dochádza k vzostupu glykémie a následne k zvýšenej sekrécii inzulínu so súčasnou supresiou glukagónu. Glukóza prijatá v potrave je dominantným zdrojom energie. Zvýšená hladina inzulínu podporuje ukladanie glukózy v pečeni vo forme glykogénu, stimuluje lipogenézu a inhibuje lipolýzu a oxidáciu mastných kyselín. Ide o fyziologický anabolický stav sprevádzaný prechodným poklesom inzulínovej senzitivity [6,7].
Postabsorpčná fáza (6–12 hodín po jedle)
Glykémia a inzulinémia postupne klesajú. Dominantným zdrojom glukózy je pečeňová glykogenolýza. S poklesom zásob glykogénu mierne stúpa glukagón a postupne sa zvyšuje podiel glukoneogenézy na produkcii glukózy. Lipolýza a betaoxidácia mastných kyselín sa začínajú zvyšovať, ketogenéza je však ešte minimálna [8,9].
Skorý pôst (12–24 hodín)
Pečeňový glykogén sa postupne vyčerpáva. Glukoneogenéza sa podieľa na produkcii glukózy približne 40–60 %. Významnú úlohu zohráva aj glukoneogenéza v obličkách, ktoré sa v tomto štádiu môžu na celkovej produkcii glukózy podieľať až 20–40 % v závislosti od individuálnych faktorov [10,11]. Zvyšuje sa lipolýza, betaoxidácia a produkcia ketolátok. Nezávisle od redukcie telesnej hmotnosti sa pozoruje aj skoré zlepšenie inzulínovej senzitivity, jeho rozsah je však individuálny [4,12].
Stredne dlhý pôst (24–72 hodín)
Pečeňový glykogén je prakticky vyčerpaný a hlavný zdroj endogénnej produkcie glukózy tvorí glukoneogenéza (približne 80 %). Významne narastá betaoxidácia mastných kyselín a ketogenéza. Mozog začína postupne zvyšovať využívanie ketolátok (približne 30–40 % energetických potrieb po 48 hod). Aktivuje sa AMPK, inhibuje sa mTOR a experimentálne dáta naznačujú zvýšenie autofágie, presná dynamika u ľudí však nie je úplne kvantifikovaná. Stúpa hodnota fibroblastového rastového faktora 21 (FGF21 – Fibroblast Growth Factor 21) – hormonálneho signálu adaptácie na hladovanie [13,14]. FGF21 je proteín produkovaný hlavne v pečeni, ktorý reguluje metabolizmus glukózy a tukov, zvyšuje citlivosť na inzulín, podporuje spaľovanie tukov a hrá úlohu pri hladovaní a energetickom strese.
Dlhý pôst (> 3 dni lačnenia)
Ketolátky sa stávajú významným zdrojom energie pre mozog (po 4–7 dňoch až 60–70 %). Hladina inzulínu zostáva nízka, hladina glukagónu relatívne zvýšená. Dochádza k čiastočnému „protein sparing“ efektu sprostredkovanému zvýšením rastového hormónu a adaptáciou CNS na ketolátky. Bazálny metabolizmus sa môže postupne znižovať. Glukoneogenéza pretrváva najmä pre erytrocyty a tkanivá závislé od glukózy.
Prolongovaný pôst (> 5 dní lačnenia)
Metabolizmus je adaptovaný na vysoký podiel oxidácie mastných kyselín a ketolátok. Strata svalových bielkovín je relatívne redukovaná v porovnaní s prvými dňami pôstu, nie však eliminovaná. Úroveň autofágie u ľudí nie je možné presne kvantifikovať, no dostupné dáta poukazujú jej zvýšenie.
Metabolický „switch“ a jeho význam
Metabolický „switch“ predstavuje postupný prechod od dominantného využívania glukózy k zvýšenej oxidácii mastných kyselín a produkcii ketolátok, ku ktorému dochádza približne po 12–18 hodinách lačnenia (individuálne variabilne). Tento proces je primárne sprostredkovaný poklesom inzulinémie pri relatívne stabilnej glykémii a vzostupom glukagónu, čo vedie k aktivácii lipolýzy a ketogenézy.
Samotný metabolický switch predstavuje fyziologickú adaptáciu na nedostatok príjmu energie. Opakované dosahovanie tohto stavu v rámci IF môže (najmä pri súčasnom energetickom deficite) prispieť k zlepšeniu metabolickej flexibility, inzulínovej senzitivity a k redukcii viscerálneho a hepatálneho tuku. Redukcia telesnej hmotnosti však nie je inherentným cieľom metabolického switchu, ale potenciálnym dôsledkom celkového energetického deficitu.
Jedným z hlavných mechanistických cieľov IF je periodické dosahovanie metabolického switchu a zlepšenie metabolickej flexibility. Redukcia telesnej hmotnosti môže byť sekundárnym dôsledkom.
Reakcia kľúčových orgánov na lačnenie
Mozog postupne prechádza z využívania glukózy na ketolátky. Po 12–24 hodinách pôstu začína ketogenéza v dôsledku vyčerpanie pečeňového glykogénu po cca 12–18 hodinách lačnenia, nízkej hladiny inzulínu a vysokej hladiny glukagónu. Vzniká nadbytok acetyl-CoA z betaoxidácie MK, ktorého vstup do Krebsovho cyklu je limitovaný nedostatkom oxaloacetátu presmerovaného na glukoneogenézu. Po 24–48 hodinách je ketogenéza už funkčne významná a po približne 2–3 dňoch ketolátky pokrývajú 30–40 % energetických potrieb mozgu, klesá potreba glukoneogenézy z bielkovín, šetria sa aminokyseliny. Pri dlhšom pôste (> 3–5 dní) sa ketolátky stávajú dominantným zdrojom energie (60–70 %) pre mozog [8,20]. Spotreba glukózy je výrazne redukovaná a šetrí sa hlavne pre erytrocyty a dreň obličiek.
Srdce po nočnom lačnení a poklese inzulinémie preferenčne zvyšuje využívanie ketolátok, pričom mastné kyseliny zostávajú trvalo významným zdrojom energie [21,22].
Kôra obličky ako hlavný zdroj energie využíva mastné kyseliny – oxiduje ich betaoxidáciou. Významne sa podieľa na glukoneogenéze, a to najmä počas hladovania. Hlavnými substrátmi renálnej glukoneogenézy sú laktát a glutamín. Po približne 48–72 hodinách pôstu môže z obličiek pochádzať až 20–25 % endogénnej produkcie glukózy [10,11]. Dreň obličky, ktorá je relatívne hypoxická, využíva predovšetkým glukózu prostredníctvom anaeróbnej glykolýzy [23].
Pečeň počas skorého pôstu uvoľňuje glukózu mechanizmom glykogenolýzy a s úbytkom glykogénu zapája glukoneogenézu. Pri dlhšom pôste dochádza k oxidácii mastných kyselín a tvorbe ketolátok, ktoré zásobujú najmä mozog a srdce [8,20,23]. Pečeň samotná ketolátky nevyužíva, ale exportuje ich ako alternatívny energetický substrát.
Hlavné metabolické účinky IF
Metabolické účinky IF možno rozdeliť do niekoľkých vzájomne prepojených oblastí.
Vplyv na energetický metabolizmus
Po približne 8–12 hodinách lačnenia dochádza k postupnému poklesu zásob pečeňového glykogénu. Aktivuje sa lipolýza v tukovom tkanive a betaoxidácia mastných kyselín (MK) v pečeni, pričom rastie produkcia ketolátok (beta-hydroxybutyrát, acetoacetát). Organizmus prechádza do tzv. metabolic switch, teda z prevažne glukózového metabolizmu na tukovo-ketónový metabolizmus [4,8]. Čas prechodu závisí od zásob glykogénu, inzulínovej senzitivity a zloženia predchádzajúcej stravy.
Homeostáza glukózy a inzulínová senzitivita
Pri IF dochádza k poklesu glykémie nalačno, zníženiu postprandiálnej glykemickej variability a k redukcii HbA1c približne o 0,25–0,91 %, najmä u osôb s inzulínovou rezistenciou alebo prediabetom [24,25]. Súčasne klesá bazálna hladina inzulínu a zvyšuje sa citlivosť periférnych tkanív na inzulín [5]. V niektorých štúdiách, najmä pri „early time-restricted feeding (eTRF)“, sa pozoruje zlepšenie inzulínovej senzitivity aj pri minimálnej zmene telesnej hmotnosti, čo naznačuje, že časovanie príjmu potravy môže mať metabolické účinky čiastočne nezávislé od redukcie hmotnosti.
Metabolizmus lipidov
IF vedie k poklesu plazmatických triglyceridov a k miernemu zlepšeniu lipidového profilu, charakterizovanému miernym poklesom LDL-cholesterolu a miernym vzostupom HDL-cholesterolu [17,26].
Hmotnosť a telesná kompozícia
Ak je IF sprevádzaný negatívnou energetickou bilanciou, dochádza k redukcii telesnej hmotnosti približne o 2–8 %, vrátane viscerálneho tuku, čo sa prejavuje zmenšením obvodu pása o 2–5 cm [27]. Zachovanie svalovej hmoty pri IF je možné pri adekvátnom prijme proteínov a silovom tréningu; dáta o nadradenosti IF oproti kontinuálnej reštrikcii sú však nejednoznačné [28].
Hepatálny tuk
V pečeni počas prerušovaného hladovania dochádza k potlačeniu de novo lipogenézy (DNL) a k zvýšeniu betaoxidácie mastných kyselín, najmä v dôsledku poklesu inzulinémie a nižšej dostupnosti sacharidov [8]. Obsah tuku v pečeni klesá už pri miernej redukcii telesnej hmotnosti, avšak zlepšenie steatózy sa pozoruje aj bez výraznej zmeny telesnej hmotnosti. Hoci je efekt závislý najmä od miery kalorického deficitu, tento nie je striktne nutnou podmienkou [18]. Pri IF, resp. TRF (Time-Restricted Feeding) dochádza k redukcii hepatálneho tuku aj bez kalorického deficitu – cez pokles inzulínu/redukciu sacharidov, zlepšenie citlivosti na inzulín, potlačenie de novo lipogenézy a zvýšenie oxidácie tukov [78–80].
V niektorých štúdiách sa pozorovala redukcia hepatálneho tuku aj bez výrazného kalorického deficitu, avšak veľkosť efektu je variabilná a závisí od celkového energetického príjmu.
Protizápalové a antioxidačné účinky. IF podľa niektorých štúdií a metaanalýz vedie k miernemu poklesu markerov chronického subklinického zápalu (CRP, IL6, TNFα, IL8), výraznejšiemu pri súčasnom úbytku tukovej hmoty. Podľa iných štúdií sa súčasne znižuje oxidačný stres a zlepšuje mitochondriálna funkcia [19,29]. U ľudí sú výsledky heterogénne, efekt je často sprostredkovaný úbytkom tuku. IF môže zlepšiť niektoré markery oxidačného stresu; pri markeroch zápalu sú výsledky heterogénne. Dôkazy o mitochondriálnej funkcii u ľudí sú limitované.
Kardiovaskulárne účinky
Pozoruje sa zníženie systolického krvného tlaku približne o 5–10 mm Hg, zlepšenie variability srdcovej frekvencie a priaznivý vplyv na endotelovú funkciu a vaskulárnu reaktivitu [30,31]. Pokles tlaku krvi (TK) je plauzibilný, ale veľkosť efektu závisí od základného TK, redukcie hmotnosti, príjmu sodíka, alkoholu, spánku. U normotenzných jednotlivcov je efekt minimálny. Vplyv na endotel je posudzovaný na základe menších a kratších štúdií.
Účinok na hormóny a reguláciu apetítu
Pri tzv. early time-restricted feeding (eTRF) je ranná hladina kortizolu a amplitúda jeho cirkadiánneho rytmu mierne zvýšená, čo sa interpretuje ako lepšie zosúladenie periférnych biologických hodín s centrálnym cirkadiánnym rytmom. V niektorých štúdiách bez známok maladaptácie je klinický význam neistý [32]. Pri IF 16 : 8 sa neočakávajú klinicky významné trvalé zmeny katecholamínov; humánne dôkazy sú však limitované [16]. Počas pôstu narastá pulzatilita sekrécie rastového hormónu (STH), najmä pri hladovaní dlhšom ako 24–48 hodín; pri dennom IF je efekt menší. Hladiny IGF1 klesajú výraznejšie až pri dlhšom hladovaní [15]. Leptín klesá už počas prvých týždňov IF a paralelne s redukciou tukovej hmoty [33]. Údaje o adiponektíne sú heterogénne; mierny vzostup sa pozoruje najmä pri dlhšom trvaní IF [34]. Pri eTRF dochádza k rannému poklesu grelínu a vzostupu PYY, čo by mohlo korelovať s lepšou reguláciou apetítu [35]. Údaje o GLP1-RA sú u ľudí však zatiaľ limitované a efekt IF je slabší než pri liečbe GLP1-RA [36].
Autofágia a prerušované lačnenie
Autofágia ako centrálny adaptačný mechanizmus pôstu
Autofágia predstavuje kľúčový mechanizmus bunkovej adaptácie na nedostatok živín a konzervovaný proces bunkovej recyklácie sprostredkovaný lyzosómami, ktorého úlohou je odstránenie poškodených organel a proteínov a udržiavanie bunkovej homeostázy. Považuje sa tiež za centrálnu regulačnú platformu prepájajúcu signálne dráhy mTOR, AMPK, sirtuínov, p53 a inflamasómu [37–39]. Je regulovaná predovšetkým prostredníctvom inhibície mTOR a aktivácie AMPK signálnej dráhy [37–39].
Autofágia pôsobí ako integračný uzol medzi metabolizmom, imunitnou odpoveďou, oxidačným stresom a bunkovým starnutím, čím vysvetľuje široké spektrum systémových účinkov IF. Väčšina klinických efektov je zatiaľ podložená iba experimentálne.
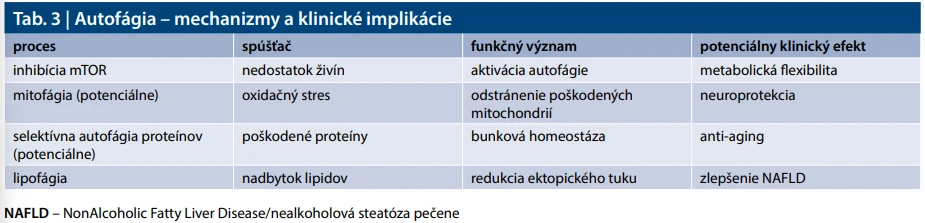
Presná časová dynamika autofágie u ľudí a jej význam nie je jednoznačne stanovená a veľa vychádza z extrapolácie. V posledných rokoch však pribúdajú aj štúdie u ľudí, ktoré naznačujú zmeny markerov autofágie (ULK1, LC3-II, p62, Beclin1, LAMP1, LAMP2, SQSTM1 a ďalšie) už po 12–24 hodinách lačnenia, pričom významnejší efekt by sa mohol prejavovať pri opakovanom alebo dlhšom pôste, avšak klinický význam týchto zmien zostáva nejasný, [11,12,37,38]. Porucha autofágie sa v experimente spája s akumuláciou poškodených proteínov, inzulínovou rezistenciou, neurodegeneráciou, imunopatológiou a metabolicky indukovaným chronickým subklinickým zápalom (tzv. metaflammation). Autofágia tlmí aktiváciu inflamasómu a produkciu IL1β a je považovaná za jeden z mechanizmov ovplyvňujúcich bunkové starnutie [40]. Autofágia je úzko prepojená s adipozitou; jej dysfunkcia vedie k poruche funkcie adipocytov a inzulínovej rezistencii. Mikrobiota moduluje autofágiu prostredníctvom krátkoreťazcových mastných kyselín (SCFA – Short Chain Fatty Acid), ktoré ovplyvňujú črevnú bariéru a subklinický zápal [41].
Časovanie a význam autofágie
Pri procese autofágie zohráva kľúčovú úlohu stav aktivácie mTOR (mechanistic Target Of Rapamycin). mTOR je hlavný bunkový senzor živín a rastových signálov, ktorý rozhoduje, či bunka bude rásť, alebo recyklovať). Aktivuje sa pri dostatku glukózy a zvýšenej hladine inzulínu a pokiaľ sú k dispozícii aminokyseliny (hlavne leucín). Aktívny mTOR blokuje autofágiu, a naopak pokles mTOR autofágiu štartuje. AMPK (AMPK – Adenosine Monophosphate-activated Protein Kinase), je senzor energetického stresu bunky. Aktivuje sa, keď klesá ATP, narastá AMP, bunka má energetický deficit. Efekt aktívnej AMPK zvyšuje oxidáciu tukov, znižuje syntézu lipidov a proteínov, aktivuje autofágiu – priamo aktivuje ULK1, inhibuje mTOR. V stave nasýtenia je mTOR zvýšený a AMPK znížená, autofágia je vypnutá, a naopak v stave hladovania je mTOR znížený a AMPK zvýšená a autofágia je zapnutá. Autofágia je teda adaptívny mechanizmus prežitia, nie systémový „detox“ organizmu.
Tieto zmeny sa výraznejšie objavia, až keď sa vyčerpá pečeňový glykogén (približne 12–24 hod) a začne ketogenéza (približne 18–30 hod). Priame dôkazy o časovaní autofágie u ľudí sú obmedzené a dostupné údaje naznačujú zvýšenie markerov po 24–36 hod pôstu.
Krátkodobý pôst (16–24 hod): naznačený nárast markerov autofágie (ULK1, LC3 konverzia, LC3-II, p62) v pečeni a kostrovom svale (> 12–16 hod – experiment, aj malé humánne štúdie), podpora lipolýzy a ketogenézy [37].
Stredne dlhý pôst (2–3 dni): na základe experimentálnych modelov výraznejšia aktivácia autofágie aj v mozgu a srdci; zvýšená sekrécia rastového hormónu (STH – SomatoTropin Hormon) chráni svalovú hmotu [15,38].
Dlhší pôst (≥ 3–5 dní): experimentálne údaje naznačujú apoptózu poškodených imunitných buniek a aktiváciu hematopoetických kmeňových buniek; humánne dôkazy sú však zatiaľ obmedzené [42].
Biologický význam autofágie pri prerušovanom lačnení
Autofágia znižuje oxidačný stres a zápal, zlepšuje inzulínovú senzitivitu a podporuje bunkové prežitie. Najpresvedčivejšie sú dôkazy o metabolických benefitoch IF (redukcia hmotnosti, zlepšenie inzulínovej senzitivity, pokles krvného tlaku) [8,19], zatiaľ čo priamy podiel autofágie na týchto účinkoch zostáva predmetom výskumu. Ďalej sa uplatňuje neuroprotekcia, kardiovaskulárna ochrana a potenciálne antiaging účinky [38,40].
Prerušované lačnenie a črevná mikrobiota
Intermitentné lačnenie (IF) ovplyvňuje aj zloženie a funkciu črevnej mikrobioty. Niektoré štúdie naznačujú, že IF zvyšuje diverzitu črevných baktérií, ktorá je považovaná za marker zdravej mikrobioty [43,44]. Dochádza k nárastu priaznivých bakteriálnych taxónov, najmä Akkermansia muciniphila, Lactobacillus a Bifidobacterium [45–47]. Naopak, IF môže viesť k redukcii prozápalových baktérií, ako sú Desulfovibrio a zástupcovia čeľade Enterobacteriaceae, ktoré sa spájajú s endotoxémiou a metabolickým zápalom [50]. Tieto prejavy boli pozorované v niektorých štúdiách, nie sú však konzistentné a sú závislé od zloženia stravy (vláknina, polyfenoly, lieky). Kauzalita je neistá.
Akkermansia muciniphila sa spája so zlepšením integrity črevnej bariéry, znížením endotoxémie a zlepšením inzulínovej senzitivity [45]. Rody Lactobacillus a Bifidobacterium produkujú krátkoreťazcové mastné kyseliny (SCFA – acetát, propionát, butyrát), ktoré sú kľúčové pre výživu kolonocytov, udržiavanie integrity črevnej bariéry, moduláciu imunitnej odpovede a tlmenie nízkostupňového zápalu (leaky gut) [46,48].
Zvýšené zastúpenie rodu Prevotella sa spája s efektívnejším metabolizmom polysacharidov a vlákniny [49].
Účinky prerušovaného lačnenia na psychiku a kognitívne funkcie
Kognitívne funkcie: Experimentálne štúdie na zvieratách a limitované humánne údaje naznačujú, že IF môže zlepšovať pamäť, učenie a pozornosť [51,52]. Zvýšená produkcia ketolátok, najmä betahydroxybutyrátu, poskytuje mozgu alternatívny, metabolicky stabilný a oxidatívne menej náročný substrát, čo môže prispievať k zvýšenej mentálnej bdelosti [53].
Neuroplasticita a neuroprotekcia: V experimentálnych modeloch IF stimuluje expresiu a uvoľňovanie mozgového neurotrofického faktora (BDNF – Brain-Derived Neurotrophic Factor), ktorý podporuje neurogenézu, synaptickú plasticitu a prežitie neurónov. Tento mechanizmus je spájaný s neuroprotekciou a potenciálnou ochranou pred neurodegeneratívnymi ochoreniami, ako sú Alzheimerova a Parkinsonova choroba, pričom dôkazy u ľudí sú zatiaľ prevažne nepriame [51,54].
Nálada a emočná stabilita: Niektoré klinické pozorovania a menšie štúdie poukazujú na zlepšenie nálady a zníženie symptómov úzkosti a depresie počas IF. Pravdepodobne ide o kombináciu hormonálnych zmien (pokles inzulínu, zvýšenie adiponektínu, stabilizácia cirkadiánneho rytmu kortizolu) a zníženia systémového zápalu [55,56].
Odolnosť voči stresu a autonómna regulácia: IF aktivuje miernu metabolickú stresovú odpoveď (hormesis), ktorá zvyšuje odolnosť mozgu voči oxidačnému a metabolickému stresu [51,57]. Zlepšuje sa regulácia autonómneho nervového systému a variabilita srdcovej frekvencie, čo môže prispievať k lepšiemu zvládaniu psychickej záťaže [58].
Spánok a cirkadiánne rytmy: Časovo obmedzené stravovanie (time-restricted eating) môže priaznivo ovplyvniť cirkadiánne rytmy. Zlepšenie kvality spánku sa pozoruje najmä pri skoršom „feeding window“ (napr. 8 : 00–16 : 00), ktoré lepšie zosúlaďuje príjem potravy s endogénnym cirkadiánnym rytmom [18,59].
Potenciálne negatívne aspekty prerušovaného lačnenia
U niektorých jedincov môže IF, najmä v adaptačnej fáze, viesť k podráždenosti, zhoršeniu nálady alebo problémom s koncentráciou. Pri nesprávnej aplikácii (extrémne dlhé hladovanie, prejedanie sa počas „feeding window“) môže IF prispievať k rozvoju alebo exacerbácii porúch príjmu potravy, čo si vyžaduje individuálny prístup a opatrnosť [60,61].
Historický a náboženský význam dobrovoľného cieleného lačnenia (pôstu)
Pôst má hlboké historické a náboženské korene naprieč civilizáciami. V Mezopotámii a starovekom Egypte bol súčasťou očistných rituálov pred náboženskými obradmi. V antickom Grécku a Ríme ho niektorí filozofi (napr. Pytagoras) odporúčali ako prostriedok sebakontroly a duševnej disciplíny; niektorí starovekí lekári považovali obmedzenie jedla za liečebný nástroj [62].
V judaizme má pôst význam pokánia a duchovnej očisty, najmä počas jom kipur – Dňa zmierenia (Lv 16 a Lv 23,27–32). Pôst je spojený s pokáním, zmierením a pokorou pred Bohom. V kresťanstve sa pôst etabloval ako príprava na veľké sviatky (napr. 40-dňový pôst pred Veľkou nocou, ktorý začína Popolcovou stredou) a je chápaný ako prostriedok pokory, pokánia, duchovnej obnovy, obrátenia srdca a posilnenia viery, vzťahu k Bohu a pripodobnenie sa Kristovi a hlbšie duchovné prežívanie tajomstva jeho obety. Biblické texty opisujú pôst tiež ako prostriedok duchovnej prípravy pred veľkou úlohou, ako bol pôst Ježiša Krista pred začiatkom verejného účinkovania (Mt 4,1–2, Hebr 10,10–14). V islame je pôst počas mesiaca Ramadán jedným z piatich pilierov viery a spája sa s duchovnou očistou a posilnením sebakontroly. V hinduizme a budhizme je pôst súčasťou duchovného tréningu, meditácie a odpútania sa od materiálneho sveta [63].
Možné klinické využitie prerušovaného lačnenia
Obezita a metabolický syndróm (MetS): IF môže viesť k redukcii telesnej hmotnosti, viscerálneho a hepatálneho tuku a k zlepšeniu inzulínovej senzitivity, krvného tlaku a lipidového profilu. Účinnosť IF je porovnateľná s kontinuálnou kalorickou reštrikciou [1–3].
Diabetes mellitus 2. typu (DM2T): IF môže zlepšiť glykemickú kontrolu a znížiť inzulínovú rezistenciu, avšak vyžaduje zvýšenú opatrnosť pri súbežnej farmakoterapii, najmä pri inzulíne a derivátoch sulfonylurey z dôvodu rizika hypoglykémie [67,68].
Kardiovaskulárna prevencia: Pozorované sú priaznivé účinky na krvný tlak, lipidové spektrum, zápalové markery a endoteliálnu funkciu. Účinky sú najvýraznejšie u pacientov s metabolickým syndrómom [5,69].
Potenciálny neuroprotektívny efekt: Experimentálne a limitované klinické údaje naznačujú zlepšenie kognitívnych funkcií a možný protektívny efekt voči neurodegeneratívnym ochoreniam, najmä prostredníctvom ketolátok, BDNF a zníženia neuroinflamácie [70,71].

Limity a riziká prerušovaného lačnenia
IF nie je vhodné pre deti a adolescentov, tehotné a dojčiace ženy, pacientov s aktívnymi poruchami príjmu potravy, pre pacientov s diabetes mellitus 1. typu, pacientov s DM2T na intenzifikovanej inzulínovej terapii, osoby s pokročilou krehkosťou (frailty), kachexiou, pre pacientov s pokročilým chronickým ochorením obličiek, pacientov po nedávnej diabetickej ketoacidóze alebo hyperosmolárnom hyperglykemickom stave (HHS – Hyperosmolar Hyperglycemic State), alebo pre neliečené alebo dekompenzované závažné komorbidity [67,72].
U diabetikov na farmakoterapii existuje zvýšené riziko hypoglykémie, najmä pri užívaní inzulínu a sulfonylurey. U niektorých pacientov môže IF viesť k prejedaniu sa počas „feeding window“ [68,73].
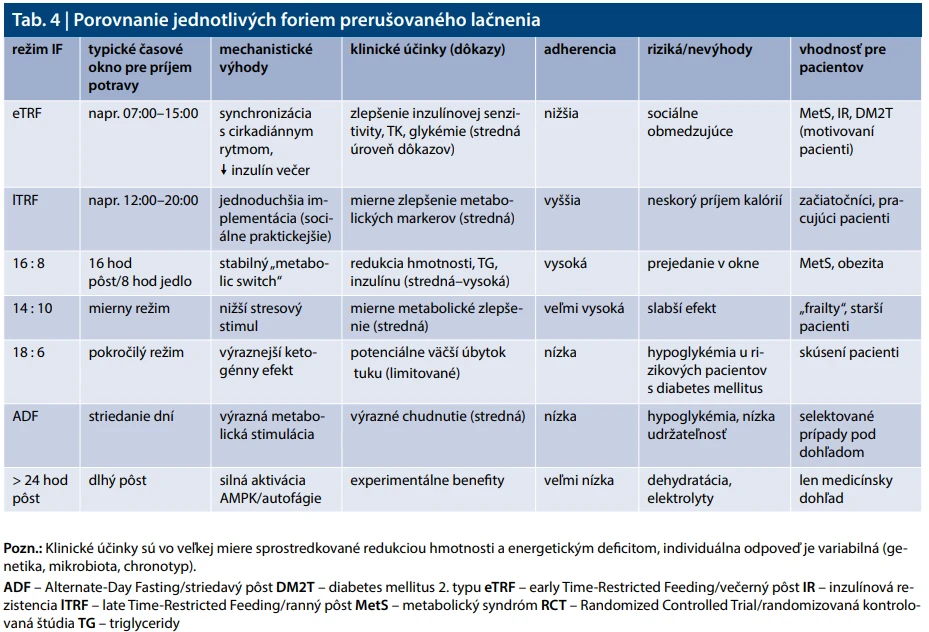
Vhodné formy prerušovaného lačnenia pre pacientov s metabolickým syndrómom a DM2T
IF 16 : 8 (5–6 dní/týždeň): Lačnenie 16 hodín (napr. od 15 : 00 do 07 : 00 hod nasledujúceho dňa – vynechá sa večera), príjem potravy počas 8-hodinového „okna“, t. j. od 7 : 00 do 15 : 00 hod. Inou možnosťou je lačnenie od 19 : 00 do 11 : 00 hod nasledujúceho dňa (vynechávajú sa raňajky) a príjem potravy od 11 : 00 do 19 : 00 hod. Ide o najjednoduchšiu a najudržateľnejšiu formu IF, vhodnú aj pre začiatočníkov. Zlepšuje glykémiu, inzulínovú senzitivitu, krvný tlak a redukuje viscerálny a hepatálny tuk [64,65].
IF 14 : 10 Miernejší režim vhodný pri vyššom riziku hypoglykémie, krehkosti alebo náročnom pracovnom režime. Stále podporuje „metabolic switch“ (glukóza tuk) [74].
IF 18 : 6 Prísnejšia forma, vhodná len pre stabilných a skúsených pacientov bez rizikovej farmakoterapie.
Pacienti s diabetes mellitus by sa mali vyhýbať forme striedavého pôstu (ADF – Alternate-Day Fasting), teda veľmi krátkym oknám pre príjem potravy (4–6 hod), obzvlášť ak sú liečení inzulínom alebo SU. Mali by sa tiež vyhýbať dlhým pôstom (> 24 hod) bez lekárskeho dohľadu. Tieto formy sú spojené s vyšším rizikom hypoglykémie, dehydratácie a nízkej udržateľnosti [68,75].
Záver
Dostupné klinické dôkazy naznačujú, že intermitentné lačnenie (IF) vrátane tzv. time-restricted eating môže u vybraných pacientov zlepšiť niektoré parametre metabolického zdravia (telesnú hmotnosť, glykémiu/inzulínovú senzitivitu a časť kardiometabolických rizikových markerov), pričom účinok často súvisí aj s energetickým deficitom, kvalitou stravy a cirkadiánnym načasovaním jedál. Údaje o znížení „subklinického“ zápalu sú heterogénne (zmeny niektorých biomarkerov sa pozorujú, no nie konzistentne naprieč štúdiami). IF môže navodiť fyziologický metabolický „switch“ smerom k vyššej oxidácii tukov/ketóze, avšak zachovanie svalovej hmoty nie je automatické a vyžaduje adekvátny príjem proteínu a silový tréning. Mechanisticky, IF aktivuje dráhy spojené so stresovou adaptáciou a autofágiou, no priame klinické dôkazy o tkanivovej autofágii u ľudí sú obmedzené. Údaje o neuroprotekcii, nálade a spánku sú sľubné, ale zatiaľ nejednoznačné a výrazne individuálne. IF nie je univerzálne riešenie: výber protokolu, cieľov a bezpečnostných opatrení má byť individualizovaný, so zvýšenou opatrnosťou a často aj s medicínskym dohľadom u rizikových skupín (najmä pri antidiabetickej liečbe s rizikom hypoglykémie, v gravidite, pri poruchách príjmu potravy a u krehkých pacientov s podhmotnosťou) [29,64,70].
Sources
-
Reaven GM. Banting lecture 1988. Role Role of insulin resistance in human disease. Diabetes 1988; 37(12): 1595–1607. Dostupné z DOI: <http://dx.doi.org/10.2337/diab.37.12.1595>.
-
Shulman GI. Ectopic fat in insulin resistance, dyslipidemia, and cardiometabolic disease. N Engl J Med 2014; 371(12): 1131–1141. Dostupné z DOI: <http://dx.doi.org/10.1056/NEJMra1011035>. Erratum in N Engl J Med 2014; 371(23): 2241.
-
Hotamisligil GS. Inflammation and metabolic disorders. Nature 2006; 444(7121): 860–867.Dostupné z DOI: <http://dx.doi.org/10.1038/nature05485>.
-
Anton SD, Moehl K, Donahoo WT et al. Flipping the metabolic switch: understanding and applying the health benefits of fasting. Obesity (Silver Spring) 2018; 26(2): 254–268. Dostupné z DOI: <http://dx.doi.org/10.1002/oby.22065>.
-
Longo VD, Panda S. Fasting, circadian rhythms, and metabolism. Cell Metab 2016; 23(5): 1048–1059. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cmet.2016.05.001>.
-
Guyton AC, Hall JE. Guyton and Hall Textbook of Medical Physiology. 14th ed. Elsevier: Philadelphia, PA 2020. ISBN 978–0323597128.
-
Berg JM, Tymoczko JL, Stryer L. Biochemistry. 9th ed. Freeman: New York, NY 2021. ISBN 978–1319114671.
-
Cahill GF Jr. Fuel metabolism in starvation. Annu Rev Nutr 2006; 26 : 1–22. Dostupné z DOI: <http://dx.doi.org/10.1146/annurev.nutr.26.061505.111258>.
-
Gerich JE. Control of glucose homeostasis. Baillieres Clin Endocrinol Metab 1993; 7(3): 551–586. Dostupné z DOI: <http://dx.doi.org/10.1016/s0950–351x(05)80207–1>.
-
Gerich JE, Meyer C, Woerle HJ et al. Renal gluconeogenesis: its importance in human glucose homeostasis. Diabetes Care 2001; 24(2): 382–391. Dostupné z DOI: <http://dx.doi.org/10.2337/diacare.24.2.382>.
-
Gerich JE. Role of the kidney in normal glucose homeostasis and in the hyperglycaemia of diabetes mellitus: therapeutic implications. Diabet Med 2010; 27(2): 136-142. Dostupné z DOI: <http://doi: 10.1111/j.1464-5491.2009.02894.x>.
-
Sutton EF, Beyl R, Early KS et al. Early time-restricted feeding improves insulin sensitivity, blood pressure, and oxidative stress even without weight loss in men with prediabetes. Cell Metab 2018; 27(6): 1212–1221.e3. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cmet.2018.04.010>.
-
Inagaki T, Dutchak P, Zhao G et al. Endocrine regulation of the fasting response by PPARα-mediated induction of fibroblast growth factor 21. Cell Metab 2007; 5(6): 415–425. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cmet.2007.05.003>.
-
Fisher FM, Maratos-Flier E. Understanding the physiology of FGF21. Annu Rev Physiol 2016; 78 : 223–241. Dostupné z DOI: <http://dx.doi.org/10.1146/annurev-physiol-021115–105339>.
-
Ho KY, Veldhuis JD, Johnson ML et al. Fasting enhances growth hormone secretion and amplifies the complex rhythms of growth hormone secretion in man. J Clin Invest 1988; 81(4): 968–975. Dostupné z DOI: <http://dx.doi.org/10.1172/JCI113450>.
-
Heilbronn LK, Smith SR, Ravussin E. Alternate-day fasting in nonobese subjects: effects on body weight, body composition, and energy metabolism. Am J Clin Nutr 2005; 81(1): 69–73. Dostupné z DOI: <http://dx.doi.org/10.1093/ajcn/81.1.69>.
-
Tinsley GM, La Bounty PM. Effects of intermittent fasting on body composition and clinical health markers in humans. Nutr Rev 2015; 73(10): 661–674. Dostupné z DOI: <http://dx.doi.org/10.1093/nutrit/nuv041>.
-
Wilkinson MJ, Manoogian ENC, Zadourian A et al. Ten-hour time-restricted eating reduces weight, blood pressure, and atherogenic lipids in patients with metabolic syndrome. Cell Metab 2020; 31(1): 92–104.e5. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cmet.2019.11.004>.
-
Mattson MP, Moehl K, Ghena N et al. Intermittent metabolic switching, neuroplasticity and brain health. Nat Rev Neurosci 2018; 19(2): 63–80. Dostupné z DOI: <http://dx.doi.org/10.1038/nrn.2017.156>. Erratum in: Nat Rev Neurosci 2020; 21(8): 445. Dostupné z DOI: <http://dx.doi.org/10.1038/s41583–020–0342-y>.
-
Hall JE. Guyton and Hall Textbook of Medical Physiology. 15th ed. Elsevier: Philadelphia, PA 2025. ISBN 978–0443111013.
-
Stanley WC, Recchia FA, Lopaschuk GD. Myocardial substrate metabolism in the normal and failing heart. Physiol Rev 2005; 85(3): 1093–1129. Dostupné z DOI: <http://dx.doi.org/10.1152/physrev.00006.2004>.
-
Taegtmeyer H, Sen S, Vela D. Return to the fetal gene program: a suggested metabolic link to gene expression in the heart. Ann N Y Acad Sci 2010; 1188 : 191–198. Dostupné z DOI: <http://doi: 10.1111/j.1749-6632.2009.05100.x>.
-
Koeppen BM, Stanton BA. Renal Physiology. 6th ed. Mosby/Elsevier: St Louis, MO 2021. ISBN 978–0323722105.
-
Carter S, Clifton PM, Keogh JB. The effects of intermittent compared to continuous energy restriction on glycaemic control in type 2 diabetes; a pragmatic pilot trial. Diabetes Research and Clinical Practice 2016; 122 : 106–112. Dostupné z DOI: <https://doi.org/10.1016/j.diabres.2016.10.010>.
-
Patterson RE, Sears DD. Metabolic effects of intermittent fasting. Annu Rev Nutr 2017; 37 : 371–393. Dostupné z DOI: <http://dx.doi.org/10.1146/annurev-nutr-071816–064634>.
-
Varady KA. Intermittent versus daily calorie restriction: which diet regimen is more effective for weight loss? Obes Rev 2011; 12(7): e593–601. Dostupné z DOI: <http://dx.doi.org/10.1111/j.1467–789X.2011.00873.x>.
-
Harris L, Hamilton s, Azevedo LA et al. Intermittent fasting interventions for treatment of overweight and obesity in adults: a systematic review and meta-analysis. V JBI Database of Systematic Reviews and Implementation Reports 2018; 16(2): 507–547. Dostupné z DOI: <http://doi: 10.11124/JBISRIR-2016-003248>.
-
Moro T, Tinsley G, Bianco A et al. Effects of eight weeks of time-restricted feeding (16/8) on basal metabolism, maximal strength, body composition, inflammation and cardiovascular risk in resistance-trained males. J Transl Med 2016; 14(1): 290. Dostupné z DOI: <http://dx.doi.org/10.1186/s12967–016–1044–0>.
-
de Cabo R, Mattson MP. Effects of intermittent fasting on health, aging, and disease. N Engl J Med 2019; 381(26): 2541–2551. Dostupné z DOI: <http://dx.doi.org/10.1056/NEJMra1905136>. Erratum in: N Engl J Med 2020; 382(3): 298. Dostupné z DOI: <http://dx.doi.org/10.1056/NEJMx190038>. Erratum in: N Engl J Med 2020; 382(10): 978. Dostupné z DOI: <http://dx.doi.org/10.1056/NEJMx200002>.
-
Sutton EF, Beyl R, Early KS et al. Early time-restricted feeding improves insulin sensitivity, blood pressure, and oxidative stress even without weight loss in men with prediabetes. Cell Metab 2018; 27(6): 1212–1221.e3. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cmet.2018.04.010>.
-
Regmi P, Heilbronn LK. Time-restricted eating: benefits, mechanisms, and challenges in translation. iScience 2020; 23(6): 101161. Dostupné z DOI: <http://dx.doi.org/10.1016/j.isci.2020.101161>.
-
Wehrens SM, Hampton SM, Kerkhofs M et sl. Mood, alertness, and performance in response to sleep deprivation and recovery sleep in experienced shiftworkers versus non-shiftworkers. Chronobiol Int 2012; 29(5): 537–548. Dostupné z DOI: <http://doi: 10.3109/07420528.2012.675258>.
-
Harvie MN, Pegington M, Mattson MP et al. The effects of intermittent or continuous energy restriction on weight loss and metabolic disease risk markers. Int J Obes (Lond) 2011; 35(5): 714–727. Dostupné z DOI: <http://dx.doi.org/10.1038/ijo.2010.171>.
-
Cho Y, Hong N, Kim KW et al. The effectiveness of intermittent fasting to reduce body mass index and glucose metabolism: a systematic review and meta-analysis. J Clin Med 2019; 8(10): 1645. Dostupné z DOI: <http://dx.doi.org/10.3390/jcm8101645>.
-
Jamshed H, Beyl RA, Della Manna DL et al. Early Time-Restricted Feeding Improves 24-Hour Glucose Levels and Affects Markers of the Circadian Clock, Aging, and Autophagy in Humans. Nutrients 2019; 11(6): 1234. Dostupné z DOI: <http://doi: 10.3390/nu11061234>.
-
Nauck MA, Meier JJ. Incretin hormones: their role in health and disease. Diabetes Obes Metab 2018; 20(Suppl 1): 5–21. Dostupné z DOI: <http://dx.doi.org/10.1111/dom.13129>.
-
Alirezaei M, Kemball CC, Flynn CT et al. Short-term fasting induces profound neuronal autophagy. Autophagy 2010; 6(6):702–710. Dostupné z DOI: <http://dx.doi.org/10.4161/auto.6.6.12376>.
-
Mizushima N, Levine B. Autophagy in human diseases. N Engl J Med 2020; 383(16): 1564–1576. Dostupné z DOI: <http://dx.doi.org/10.1056/NEJMra2022774>.
-
Galluzzi L, Baehrecke EH, Ballabio A et al. Molecular definitions of autophagy and related processes. EMBO J 2017; 36(13): 1811–1836. Dostupné z DOI: <http://dx.doi.org/10.15252/embj.201796697>.
-
Levine B, Kroemer G. Biological functions of autophagy genes: a disease perspective. Cell 2019; 176(1–2): 11–42. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cell.2018.09.048>.
-
Koh A, De Vadder F, Kovatcheva-Datchary P et al. From dietary fiber to host physiology: short-chain fatty acids as key bacterial metabolites. Cell 2016; 165(6): 1332–1345. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cell.2016.05.041>.
-
Cheng CW, Adams GB, Perin L et al. Prolonged fasting reduces IGF-1/PKA to promote hematopoietic-stem-cell-based regeneration. Cell Stem Cell 2014; 14(6): 810–823. Dostupné z DOI: <http://dx.doi.org/10.1016/j.stem.2014.04.014>.
-
Sonnenburg JL, Bäckhed F. Diet–microbiota interactions as moderators of human metabolism. Nature 2016; 535(7610): 56–64. Dostupné z DOI: <http://dx.doi.org/10.1038/nature18846>.
-
Zmora N, Suez J, Elinav E. You are what you eat: diet, health and the gut microbiota. Nat Rev Gastroenterol Hepatol 2019; 16(1): 35–56. Dostupné z DOI: <http://dx.doi.org/10.1038/s41575–018–0061–2>.
-
Everard A, Belzer C, Geurts L et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA 2013; 110(22): 9066–9071. Dostupné z DOI: <http://dx.doi.org/10.1073/pnas.1219451110>.
-
Ríos-Covián D, Ruas-Madiedo P, Margolles A et al. Intestinal short chain fatty acids and their link with diet and human health. Front Microbiol 2016; 7 : 185. Dostupné z DOI: <http://dx.doi.org/10.3389/fmicb.2016.00185>.
-
Dao MC, Everard A, Aron-Wisnewsky J et al. Akkermansia muciniphila and improved metabolic health during dietary intervention in obesity. Gut 2016; 65(3): 426–436. Dostupné z DOI: <http://dx.doi.org/10.1136/gutjnl-2014–308778>.
-
Kovatcheva-Datchary P, Nilsson A, Akrami R. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab 2015; 22(6): 971–982. Dostupné z DOI: <http://doi: 10.1016/j.cmet.2015.10.001>.
-
Kovatcheva-Datchary P, Nilsson A, Akrami R et al. Dietary fiber-induced improvement in glucose metabolism is associated with increased abundance of Prevotella. Cell Metab 2015; 22(6): 971–982. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cmet.2015.10.001>.
-
Cani PD, Amar J, Iglesias MA, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007; 56(7): 1761–1772. Dostupné z DOI: <http://dx.doi.org/10.2337/db06–1491>.
-
Mattson MP, Moehl K, Ghena N et al. Intermittent metabolic switching, neuroplasticity and brain health. Nat Rev Neurosci 2018;19(2): 81–94. Dostupné z DOI: <http://dx.doi.org/10.1038/nrn.2017.156>. Erratum in: Nat Rev Neurosci 2020; 21(8): 445. Dostupné z DOI: <http://dx.doi.org/10.1038/s41583–020–0342-y>.
-
Witte AV, Fobker M, Gellner R et al. Caloric restriction improves memory in elderly humans. Proc Natl Acad Sci USA 2009; 106(4): 1255–1260. Dostupné z DOI: <http://dx.doi.org/10.1073/pnas.0808587106>.
-
Newman JC, Verdin E. β-hydroxybutyrate: a signaling metabolite. Annu Rev Nutr 2017; 37 : 51–76. Dostupné z DOI: <http://dx.doi.org/10.1146/annurev-nutr-071816–064916>.
-
Marosi K, Mattson MP. BDNF mediates adaptive brain and body responses to energetic challenges. Trends Endocrinol Metab 2014; 25(2): 89–98. Dostupné z DOI: <http://dx.doi.org/10.1016/j.tem.2013.10.006>.
-
Harvie MN, Pegington M, Mattson MP et al. The effects of intermittent or continuous energy restriction on weight loss and metabolic disease risk markers: a randomized trial in young overweight women. Int J Obes (Lond) 2011; 35(5): 714–727. Dostupné z DOI: <http://doi: 10.1038/ijo.2010.171>.
-
Lopresti AL, Hood SD, Drummond PD. A review of lifestyle factors that contribute to important pathways associated with major depression. J Affect Disord 2013; 148(1): 12–27. Dostupné z DOI: <http://dx.doi.org/10.1016/j.jad.2013.01.014>.
-
Calabrese EJ. Hormesis: principles and applications. Homeopathy 2015; 104(2): 69–82. Dostupné z DOI: <http://dx.doi.org/10.1016/j.homp.2015.02.007>.
-
Mager DE, Wan R, Brown M et al. Caloric restriction and intermittent fasting alter spectral measures of heart rate and blood pressure variability in rats. FASEB J 2006; 20(6): 631–637. Dostupné z DOI: <http://dx.doi.org/10.1096/fj.05–5263com>.
-
Panda S. Circadian physiology of metabolism. Science 2016; 354(6315): 1008–1015. Dostupné z DOI: <http://dx.doi.org/10.1126/science.aah4967>.
-
Tinsley GM, La Bounty PM. Effects of intermittent fasting on body composition and clinical health markersin humans. Nutr Rev 2015; 73(10): 661–674. Dostupné z DOI: <http://dx.doi.org/10.1093/nutrit/nuv041>.
-
Treasure J, Duarte TA, Schmidt U. Eating disorders. Lancet 2020; 395(10227): 899–911. Dostupné z DOI: <http://dx.doi.org/10.1016/S0140–6736(20)30059–3>.
-
Nutton V. Ancient Medicine (Sciences of Antiquity) . 2nd ed. Routledge: London 2012. ISBN 978–0415520959>.
-
Longo VD, Anderson RM. Nutrition, longevity and disease: from molecular mechanisms to interventions. Cell 2022; 185(9): 1455–1470. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cell.2022.04.002>.
-
Anton SD, Moehl K, Donahoo WT et al. Flipping the metabolic switch. Obesity (Silver Spring) 2018; 26(2): 254–268. Dostupné z DOI: <http://dx.doi.org/10.1002/oby.22065>.
-
Wilkinson MJ, Manoogian ENC, Zadourian A et al. Ten-Hour Time-Restricted Eating Reduces Weight, Blood Pressure, and Atherogenic Lipids in Patients with Metabolic Syndrome. Cell Metab 2020; 31(1): 92–104. e5. Dostupné z DOI: <http://doi: 10.1016/j.cmet.2019.11.004>.
-
Cho Y, Hong N, Kim KW et al. Effect of Time-Restricted Eating on Weight Loss and Other Metabolic Parameters in Overweight and Obese Adults Nutrients 2019; 11(6): 1387. Dostupné z DOI: <http://doi: 10.3390/nu11061387>.
-
American Diabetes Association. Standards of Care in Diabetes – 2024. Diabetes Care 2024; 47(Suppl 1): S1–S350. Dostupné z DOI: <http://dx.doi.org/10.2337/dc24-SINT>.
-
Abdelrahim D, Faris ME, Hassanein M. Impact of Ramadan Diurnal Intermittent Fasting on Hypoglycemic Events in Patients With Type 2 Diabetes: A Systematic Review of Randomized Controlled Trials and Observational Studies. Front Endocrinol (Lausanne) 2021; 12 : 624423. Dostupné z DOI: <http://doi: 10.3389/fendo.2021.624423>.
-
Sutton EF, Beyl R, Early KS, et al. Early time-restricted feeding improves metabolic health. Cell Metab 2018; 27(6): 1212–1221.e3. Dostupné z DOI: <http://dx.doi.org/10.1016/j.cmet.2018.04.010>.
-
de Cabo R, Mattson MP. Effects of Intermittent Fasting on Health, Aging, and Disease. N Engl J Med 2019; 381(26): 2541–2551. Dostupné z DOI: <http://doi:10.1056/NEJMra1905136>.
-
Newman JC, Verdin E. Ketone bodies as signaling metabolites. Trends Endocrinol Metab 2014; 25(1): 42–52. Dostupné z DOI: <http://dx.doi.org/10.1016/j.tem.2013.09.002>.
-
Volkert D, Beck AM, Cederholm T et al. ESPEN guideline on clinical nutrition and hydration in geriatrics. Clinical Nutrition 2019; 38(1): 10–47. Dostupné z DOI: <http://doi: 10.1016/j.clnu.2018.05.024>.
-
Tinsley GM, La Bounty PM. Effects of intermittent fasting on body composition and clinical health markers in humans. Nutr Rev 2015; 73(10): 661–674. Dostupné z DOI: <http://dx.doi.org/10.1093/nutrit/nuv041>.
-
Jamshed H, Beyl RA, Della Manna DL et al. Early Time-Restricted Feeding Improves 24-Hour Glucose Levels and Affects Markers of the Circadian Clock, Aging, and Autophagy in Humans. Nutrients 2019; 11(6): 1234. Dostupné z DOI: <http://doi: 10.3390/nu11061234>.
-
Trepanowski JF, Kroeger CM, Barnosky A et al. Effect of Alternate-Day Fasting on Weight Loss, Weight Maintenance, and Cardioprotection Among Metabolically Healthy Obese Adults: A Randomized Clinical Trial. JAMA Intern Med 2017; 177(7): 930–938. Dostupné z DOI: <http://dx.doi.org/10.1001/jamainternmed.2017.0936>.
-
Wei M, Brandhorst S, Shelehchi M et al. Fasting-mimicking diet and markers/risk factors for aging, diabetes, cancer, and cardiovascular disease. Sci Transl Med 2017; 9(377): eaai8700. Dostupné z DOI: <http://doi: 10.1126/scitranslmed.aai8700>.
-
Mehanna HM, Moledina J, Travis J. Refeeding syndrome: what it is, and how to prevent and treat it. BMJ 2008; 336(7659): 1495–1498. Dostupné z DOI: <http://dx.doi.org/10.1136/bmj.a301>.
-
Yin C, Li Z, Xiang Y et al. Effect of Intermittent Fasting on Non-Alcoholic Fatty Liver Disease: Systematic Review and Meta-Analysis. Front Nutr 2021; 8 : 709683. Dostupné z DOI: <http://dx.doi.org/10.3389/fnut.2021.709683>.
-
Lange M, Nadkarni D, Martin L et al. Intermittent fasting improves hepatic end points in nonalcoholic fatty liver disease: A systematic review and meta-analysis. Hepatol Commun 2023; 7(8): e0212. Dostupné z DOI: <http://dx.doi.org/10.1097/HC9.0000000000000212>.
-
Memel ZN, Wang J, Corey KE. Intermittent Fasting as a Treatment for Nonalcoholic Fatty Liver Disease: What Is the Evidence? Clin Liver Dis (Hoboken) 2022; 19(3): 101–105. Dostupné z DOI: <http://dx.doi.org/10.1002/cld.1172>.
Labels
Diabetology Endocrinology Internal medicineArticle was published in
Forum Diabetologicum

2026 Issue 1
Most read in this issue
- Moderná diabetológia sa mení na medicínu komplexných biologických súvislostí
- Pleiotropné účinky inhibítorov SGLT2 a ich podstata
- Patogénne dráhy orgánového poškodenia pri metabolickom syndróme
- Klinické postupy manažmentu prevencie vzniku cholelitiázy u vysoko rizikových pacientov liečených receptorovými agonistami GLP1